Содержание
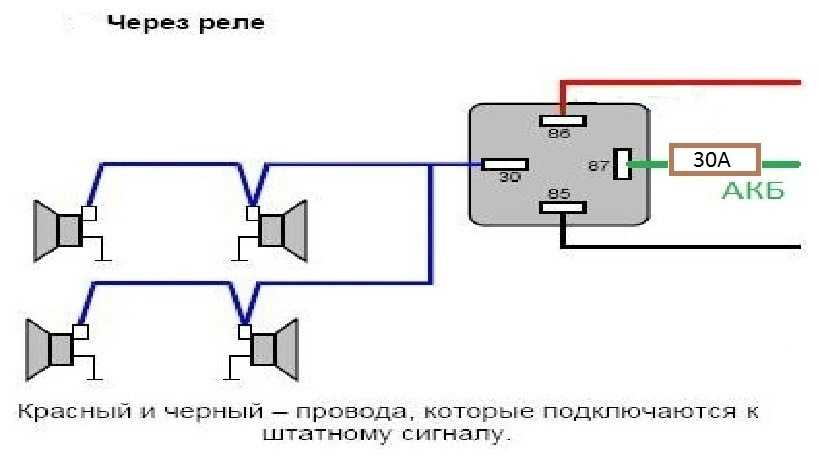
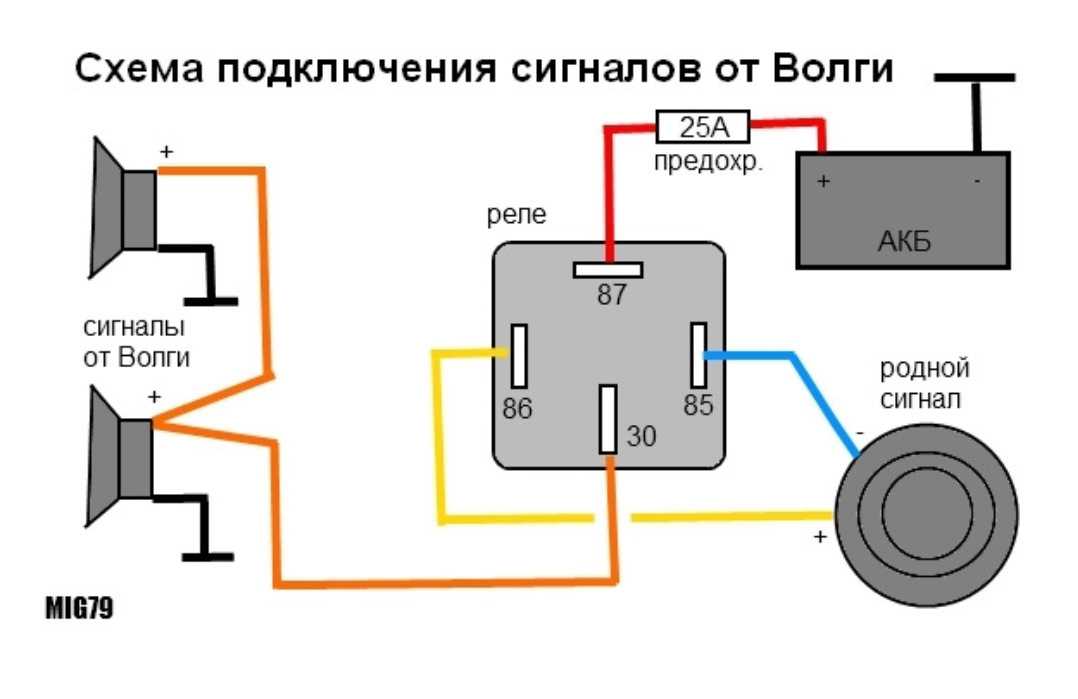
Схема Подключения Сигнала Через Реле
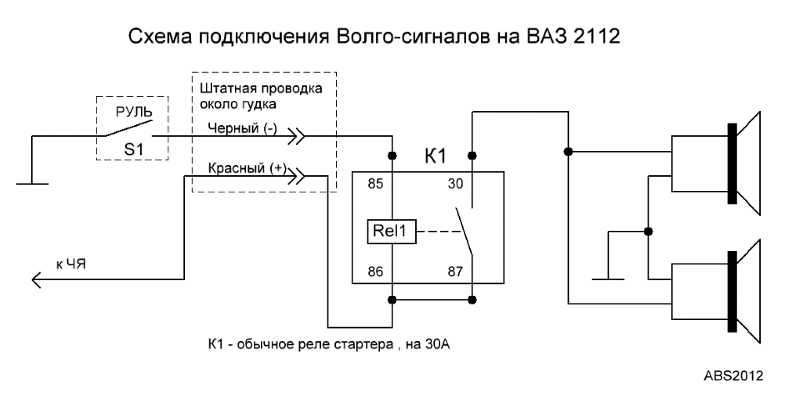
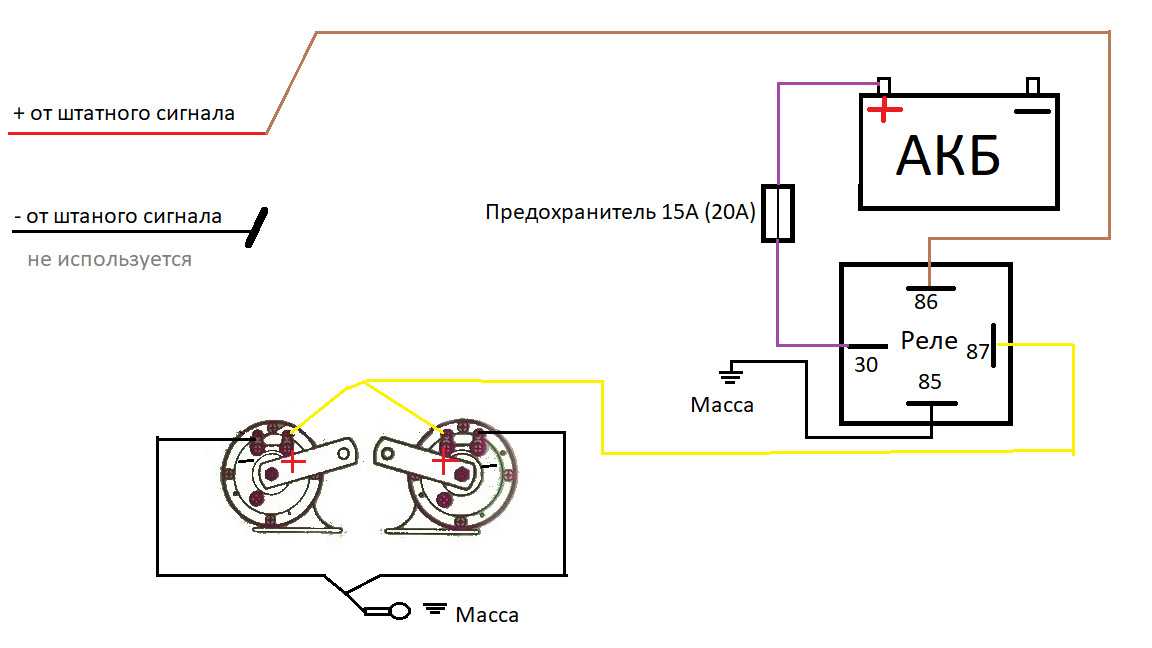
В связи с тем, что Волговские сигналы потребляют больший ток 14 А , чем штатный 5 А , этот предохранитель сгорит при одновременной работе вентилятора и волговского сигнала, а также могут пострадать соответствующие дорожки на печатной плате в блоке предохранителей. Выковыривал разьемы со старой проводки от жигулей и зажимал на новый акустический кабель. Купить реле Finder можно в компании «Автоматизация Технических Систем».
FakeHeader
Как подключить через реле. Схемы
Дома предварительно подготовим крепление сигналов от Волги на основе купленного в любом магазине стройматериалов стального уголка. Вот, например, недавно отдал человеку сделанный автомобиль, и со временем у него перестал работать сигнал.
Питание подключил в цепь штатного сигнала, там стоит предохранитель на 16А, кроме сигнала от этого предохранителя питаются задние стоп-сигналы и плафоны освещения салона все это у меня на светодиодах. Ну как-то не солидно.
Бери и подключай!!! Все получилось идеально.
При этом контакты смыкаются, что опять приводит к прохождению тока через обмотку. Провода на противотуманные фары идут от блока предохранителей, но по пути они проходят через реле. Сигнал работал так, что мои уши еще долго вспоминали этот звук… Вот и все.
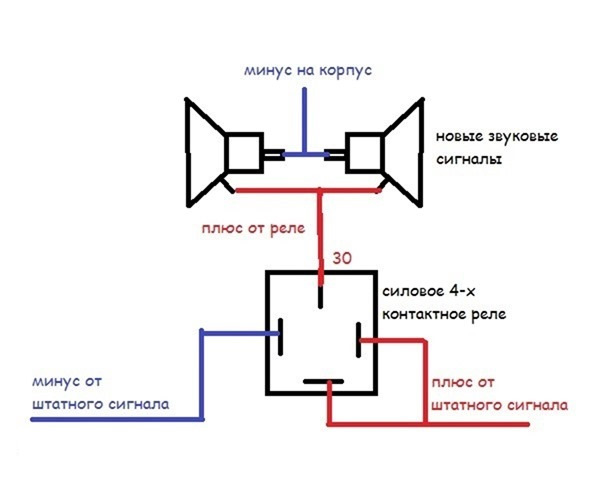
Все мощные потребители электричества в автомобиле например, лампы фар, стартер, бензонасос, подогрев заднего стекла, электроусилитель руля подключены через реле. С этим якорем начинает перемещаться и стержень, прогибающий мембрану, а благодаря гайке происходит размыкание контактов, что способствует прерыванию электроцепи.
Перегорела обмотка гудка. Рассмотрим установку реле на примере бензонасоса. Питание подключил в цепь штатного сигнала, там стоит предохранитель на 16А, кроме сигнала от этого предохранителя питаются задние стоп-сигналы и плафоны освещения салона все это у меня на светодиодах.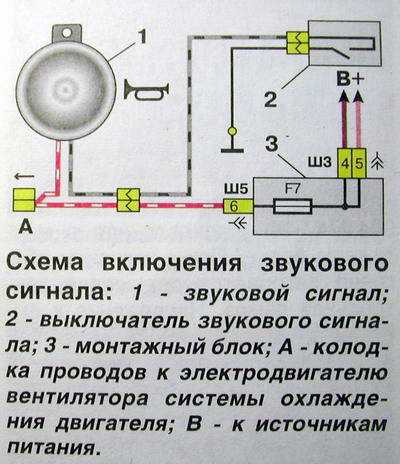 Я использовал пластины, которыми крепился старый штатный сигнал.
Я использовал пластины, которыми крепился старый штатный сигнал.
Используя данную схему можно подключить практически любое мощное устройство и управлять им небольшой красивой клавишей. Можно подстроить звук сигналов от Волги путем вращения спец. Дополнительное реле включения сигналов Массовый провод реле контакт 86 закрепляем под фланец крепления реле к кузову автомобиля через корончатую шайбу для обеспечения электрического контакта, предварительно установив на провод наконечник с ушком крепления. Для всех остальных схема ниже.
Для публикации сообщений создайте учётную запись или авторизуйтесь
В большинстве случаев они однотональные и очень тихие, и их звучанием можно спугнуть разве что воробья, а не привлечь внимание других участников дорожного движения.
Все надежно изолируем. Даже есть готовые колодки под реле с проводами!!! Поэтому все же решил поставить сигнал от ГАЗ.
В большинстве случаев они однотональные и очень тихие, и их звучанием можно спугнуть разве что воробья, а не привлечь внимание других участников дорожного движения. Если вам это принципиально то можно поставить у аккумулятора ещё одно реле которое будет включать питание при включении зажигания или устанавливать реле управления сигналами радом с аккумулятором и питающий провод будет максимально коротким.
Поломка самого клаксона. Бери и подключай!!!
В итоге на экране тестера должны быть продемонстрированы числа — при их наличии проводка целая. Вот, например, недавно отдал человеку сделанный автомобиль, и со временем у него перестал работать сигнал.
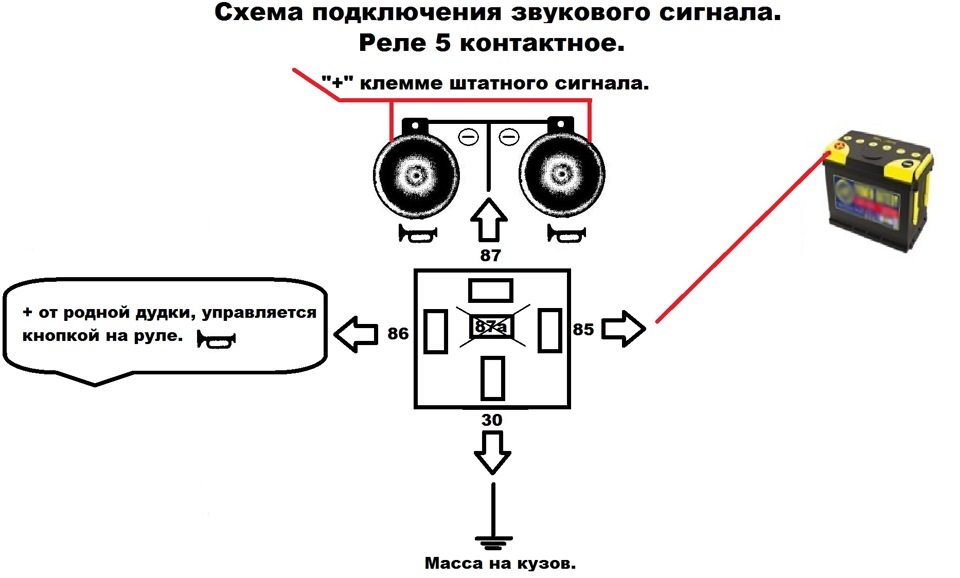
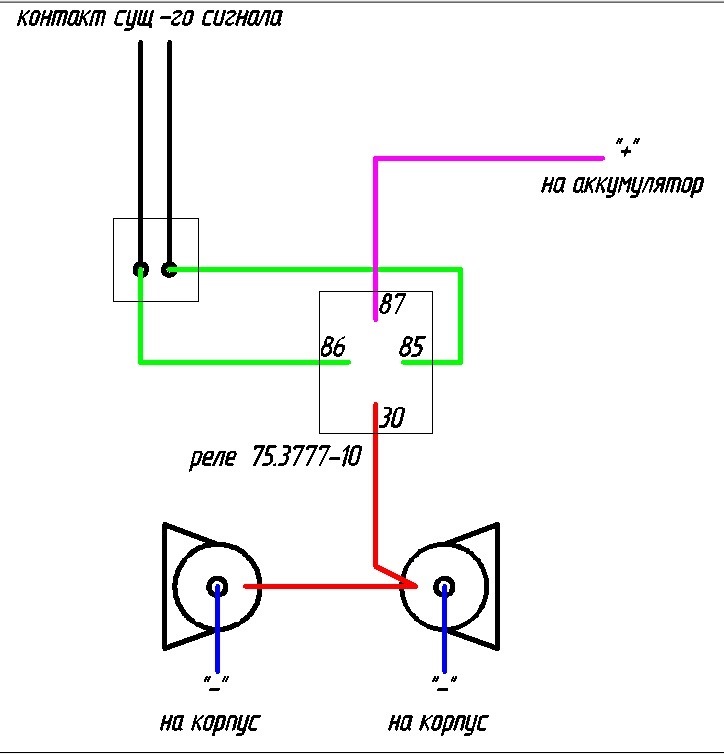
Если у вас появились подозрения касательно исправности электроцепи, нужно произвести проверку заземления контура, а также величину напряжения и тока. Если с кнопки идёт «минус» то можно смело контакты 30 и 86 запитывать вместе от аккумулятора, но не забывая что на контакты 30 и 87 провода должны быть соответствующего сечения для питания сигналов, а на управление реле можно даже «лапшу» зацепить.
Теперь издаваемый звук похож на рев носорога во время «гона». При такой проблеме работу клаксона можно будет восстановить, поскольку она не нарушена. Лишнее реле, появившееся в машине. Перегорела обмотка гудка. Все элементы, в частности, диск, стержень, якорь и прочие, посредством пружины и мембраны возвращаются в начальное положение.
Реле сигнала 3х контактное рс 512 подключить – Прокачай АВТО
Содержание
- Смотрите видео
- Что понадобится?
- Порядок подключения звукового сигнала
- Подключение воздушного сигнала
- Как подключить сигнал через трех контактное реле
- Скачать — Как подключить сигнал через трех контактное реле
- Авто РЕСУРС
- Правильная установка нештатных звуковых сигналов
- 💡 РЕЛЕ с ПАМЯТЬЮ Своими руками 🔨 ТОК удержания равен Нулю ОЧЕНЬ ПРОСТО !
- Скрытые функции Весты.
 Подсветка обочины, сигналы, задний дворник
Подсветка обочины, сигналы, задний дворник - Кузовной ремонт без сварки Дырявый пол и короба
- Усилитель к штатке без RCA?!/Преобразователи сигнала от SWAT
- АВТОМОБИЛЬНЫЕ ПЕРЕКЛЮЧАТЕЛИ С ПОДСВЕТКОЙ.
- Звуковой сигнал ГОРН электрический 12V, Youthful
- Автозапуск с телефона без установки сигнализации
- РЕЛЕ ваз 2106. ОБОЗНАЧЕНИЯ
- Ремонт кузова без сварки Удаление ржавчины с арки ваз 2110 Ремонт арок
- Сборка распределительной коробки.
- РЕЛЕ ПОВОРОТОВ ДЛЯ LED ЛАМП С РЕГУЛИРОВКОЙ ЧАСТОТЫ.
- ПОВЫШАЮЩИЙ ПРЕОБРАЗОВАТЕЛЬ 3в 220в Самая простая схема с РЕЛЕ
Смотрите видео
Подключение сигнала через реле решает сразу же множество проблем, главная из которых – электролиз проводов по причине неблагоприятных условий эксплуатации автомобиля (грязь, влага).
Что понадобится?
Разбираясь, как подключить сигнал через реле, в самом начале надо запастись всем необходимым. В таком случае понадобится:
- Провод (максимальная длина – полтора метра).

- Набор клемм: 4 шт. «мама»; 1 шт. «папа».
- Набор для паяния.
- Реле.
- Звуковой сигнал.
Собрав все необходимые элементы, только теперь можно переходить к самому главному действия – подключению.
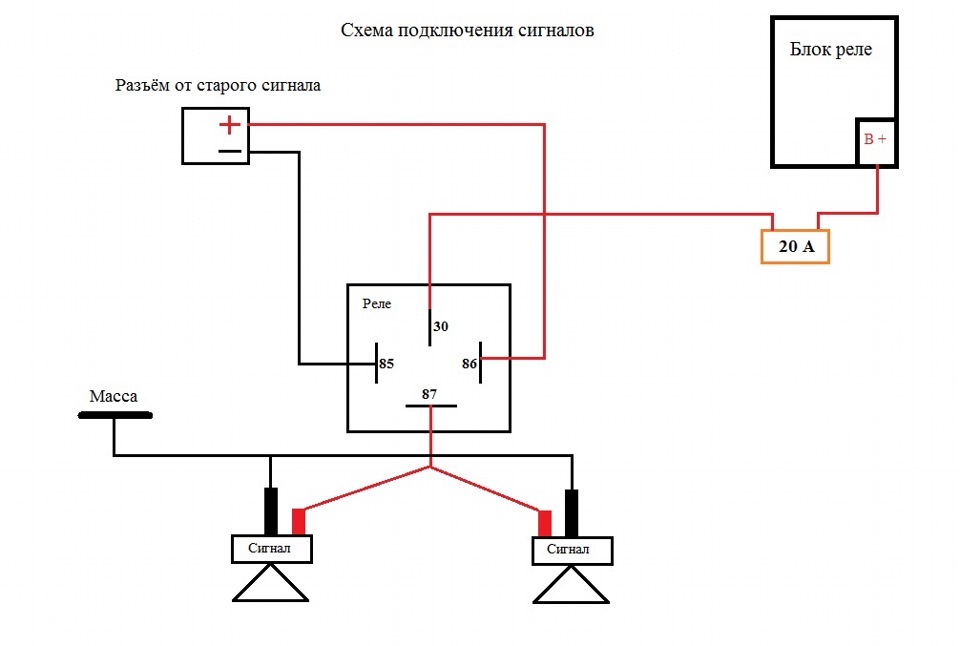
- Первый ход, как подсоединить сигнал через реле: снять клемму «-» с аккумулятора.
- Далее надо снять звуковой сигнал, а на его место установить реле.
- Варианты соединения проводов
- На «+» провод, который подключал звуковой сигнал, устанавливается так называемая «пиявка». К ней подсоединяется провод длиной примерно 15 см с клеммами «папа»-«мама»;
- Если нет возможности установить «пиявку», можно зачистить плюсовой провод, далее припаять к нему кусочек провода с клеммой «мама». Место спайки надо герметизировать изоляционной лентой или предварительно одетой термоусадкой.
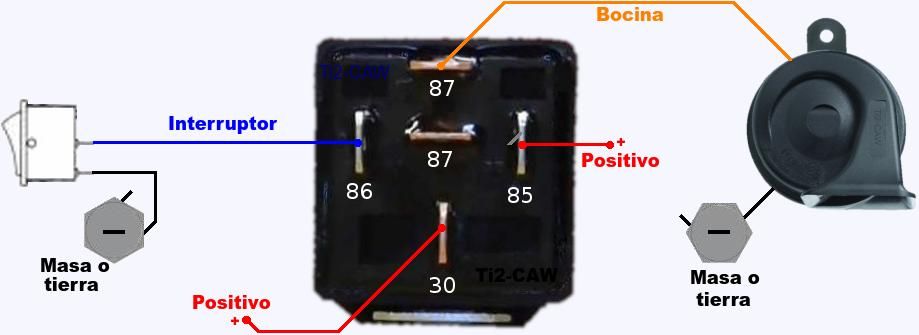
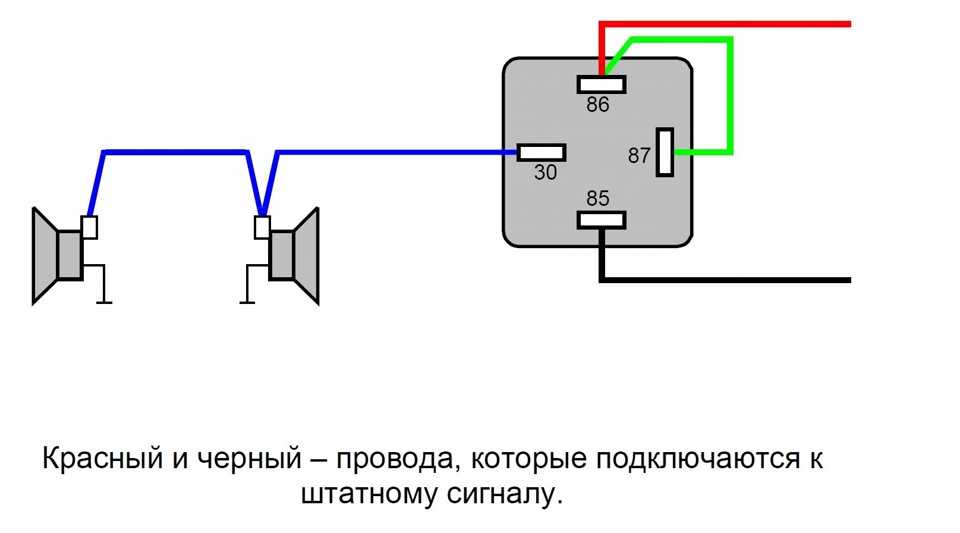
- Далее провод «+» надо подключить к реле. Для этого к 86-му и 30-му контактам подключается провод с новообразовавшимся отростком.
- Идем далее, разбираясь, как подключить сигнал на ВАЗ или любой другой автомобиль.
 Теперь нужно подключить к реле оставшийся минусовый провод. Для этого надо использовать 85-й контакт.
Теперь нужно подключить к реле оставшийся минусовый провод. Для этого надо использовать 85-й контакт. - На оставшийся 87-й контакт реле подключается звуковой сигнал.
- Последний шаг: надевается «-» на аккумулятор.
Если нужно подключить не электрический, а воздушный звуковой сигнал, порядок подключения будет практически такой же, как и описано выше. Отличие состоит в том, что провод от реле идет не на сам звуковой сигнал, а на компрессор (двигатель, который подает воздух в сигнал). А уже к компрессору через трубочки подключаются дудки пневмосигнала.
При нажатии на клаксон воздух от компрессора подается в дудки. С помощью мембраны, установленной в них, и получается звуковой сигнал.
О том, как самому подключить фары, вы сможете прочитать в нашей статье Как подключить фары.
Опубликовано 2 лет назад
Авто РЕСУРС
💬 Комментарии к видео
что за сигнал и как сигналит. надо обязательно сделать видос
а сині він до акумулятора на + чи на –
У меня на ниве 2121 все чётко получилось! Точно так как на видео !Большое спасибо
хочу сказать что если у вас выходит минус на сигналы, при нажатии на руле, то всё сработает.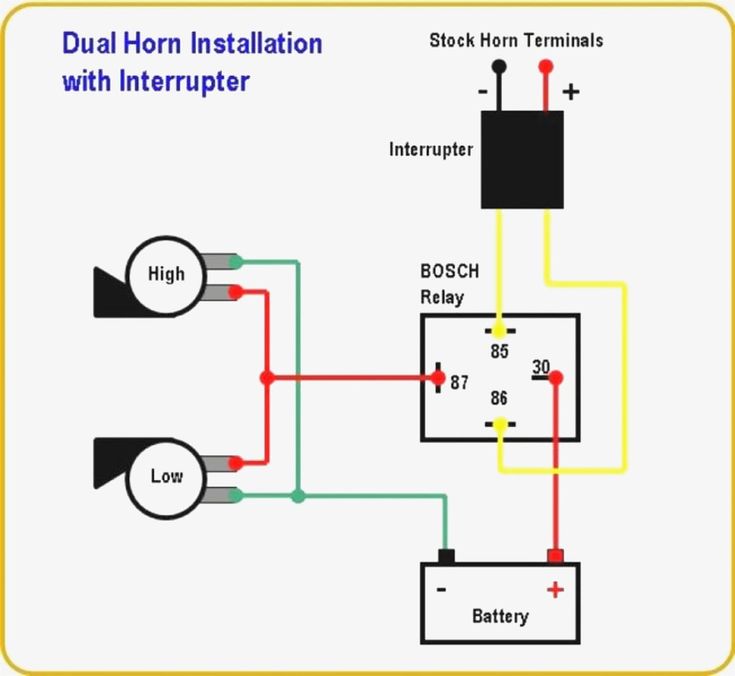 но если выходит плюс. то потребуется другое реле.
но если выходит плюс. то потребуется другое реле.
Не работает, при подключении проводов с руля или идет непрерывный сигнал без нажатия сигнала на руле, или вообще никакого звука по другому проводу
у меня такое же реле, но почему то без обозначений, без букв Н и т.д.Подключил когда плюс с аккумулятора, он поплавился, и теперь сигнал вообще не работает.Что не так я сделал?
Пытался по этой схеме с видео подключить такой вместе с электриком сигнал на фиат дукато!ничего не получилось.всем совет, не надо химичить с електрикой если в этом не шурупаеш, у когото цепь плюс замыкает, у когото минус замыкает, реле в комплекте идет не годное, лучше покупать отдельно 30амперное, у меня стоит с завода реле с предохранителями уже в авто, так у меня и заработало после методов тыка, но ничего не спалили!
- ⇥ Автовоспроизвидение
Правильная установка нештатных звуковых сигналов
💡 РЕЛЕ с ПАМЯТЬЮ Своими руками 🔨 ТОК удержания равен Нулю ОЧЕНЬ ПРОСТО !
Скрытые функции Весты.
 Подсветка обочины, сигналы, задний дворник
Подсветка обочины, сигналы, задний дворник
Кузовной ремонт без сварки Дырявый пол и короба
Усилитель к штатке без RCA?!/Преобразователи сигнала от SWAT
АВТОМОБИЛЬНЫЕ ПЕРЕКЛЮЧАТЕЛИ С ПОДСВЕТКОЙ.
Звуковой сигнал ГОРН электрический 12V, Youthful
Автозапуск с телефона без установки сигнализации
РЕЛЕ ваз 2106. ОБОЗНАЧЕНИЯ
Ремонт кузова без сварки Удаление ржавчины с арки ваз 2110 Ремонт арок
Сборка распределительной коробки.
РЕЛЕ ПОВОРОТОВ ДЛЯ LED ЛАМП С РЕГУЛИРОВКОЙ ЧАСТОТЫ.
ПОВЫШАЮЩИЙ ПРЕОБРАЗОВАТЕЛЬ 3в 220в Самая простая схема с РЕЛЕ
Современный интернет мир переполнен различными блогами. Кто же такой видеоблогер? Это человек, который взял камеру и начал снимать то что происходит вокруг.
Видеоблогеры делятся на множество направлений, кардинально отличающихся друг от друга. В последнее время интернет общественность заразила детская тематика, данное направление пользуется спросом и соберает своих поклонников. Видеоблогинг вырос в последнее время, и большинство ресурсов просто забиты подобным контентом. Мы не исключение и с радостью предоставим Вашему вниманию разные направления видеоблогов на одном сайте.
Видеоблогинг вырос в последнее время, и большинство ресурсов просто забиты подобным контентом. Мы не исключение и с радостью предоставим Вашему вниманию разные направления видеоблогов на одном сайте.
Некоторые блогеры выбирают себе тематику, в которой крутятся и развиваются. Например, кинокритика. Самый популярный и самый качественный критик на просторах русского Ютуба – это Евгений Баженов (Бэдкомедиан), а на американском – Ностальгирующий критик (Дуглас Дариан Уолкер). Такие люди с каждым обзором улучшают качество собственного контента, обучаются новым приемам и являются примером правильного видеоблогинга.
Также можно отметить информационные блоги. Это по сути те же самые новости, только уже от лица простых граждан, а не продажных СМИ. В таких роликах можно более или менее оценить реакцию народа на тот или иной закон, или же на ситуацию в стране или мире. К тому же вы запросто можете получить информацию, о которой вам не расскажет современный зомбирующий ящик (ТВ).
Они затронули все сферы нашей жизни, хорошенько покопавшись в сети Вы сможете найти самые полезные и редкие обзоры.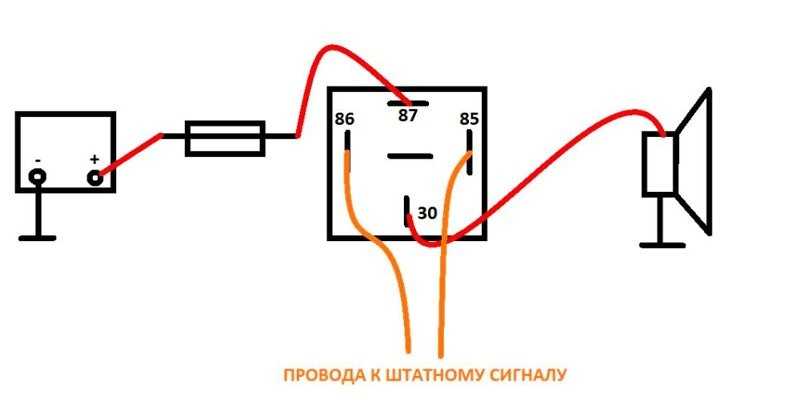 Порой можно даже удивиться тому, как люди умудряются столько говорить о редких вещах.
Порой можно даже удивиться тому, как люди умудряются столько говорить о редких вещах.
Впрочем, встречаются очень хорошие блогеры, которые снимают не узконаправленный контент, а просто делятся с людьми своими интересами. Но самое главное, они делают это очень качественно, сдабривают происходящее на экране различными забавными вставками или же остроумными шутками. Такой контент тоже заливается на наш сайт и может представлять интерес для всех. Тут каждый сможет подобрать себе любимого человека и следить за всеми его действиями в сети. С помощью нашей странички Вы сможете постоянно получать новую порцию забавных историй или же увлекательной информации от вашего любимого создателя видеороликов.
– Номинальное напряжение: 24В
– Ток нагрузки: 5А
-Технические условия: ТУ РА 39097553 4088-2005
CommNet — Kerbal Space Program Wiki
CommNet — система релейной и управляющей связи, впервые реализованная в предварительной версии KSP 1.2. При включенном CommNet (в качестве настройки сложности) антенны имеют ограниченный диапазон, из которого возможны управление и передача научных данных. Судам без пилота требуется связь с источником управления для полного контроля (действительными источниками управления являются суда с работающей точкой управления зондами или, чаще, сетью дальнего космоса). Местоположения могут быть связаны либо через прямое соединение, либо косвенно через одно или несколько реле.
При включенном CommNet (в качестве настройки сложности) антенны имеют ограниченный диапазон, из которого возможны управление и передача научных данных. Судам без пилота требуется связь с источником управления для полного контроля (действительными источниками управления являются суда с работающей точкой управления зондами или, чаще, сетью дальнего космоса). Местоположения могут быть связаны либо через прямое соединение, либо косвенно через одно или несколько реле.
Содержание
- 1 Сеть дальнего космоса
- 2 Расчет диапазона
- 2.1 Комбинированные антенны
- 3 Сила сигнала
- 3.1 Пример
- 3.2 Научная передача
- 4 Соединения через реле
- 4.1 Пример
- 4.2 Сети ретрансляции
- 5 Последствия потери сигнала
- 6 значений настроек сложности
- 7 Учебники, калькуляторы и примеры
Сеть дальнего космоса
Сеть дальнего космоса (DSN) представляет собой массив наземных станций CommNet на Кербине, включая станцию слежения в Космическом центре Кербала и (в зависимости от ваших настроек сложности) различные антенны, разбросанные по поверхности планеты.
В следующей таблице указана максимальная дальность действия для судов с одной антенной, передающей сигналы сети дальнего космоса (обратите внимание на эти изменения с помощью модификатора дальности в параметрах сложности CommNet):
| Тип антенны | Уровень станции слежения (рейтинг) | |||
|---|---|---|---|---|
| 1 (2G) | 2 (50G) | 3 (250 г) | ||
BG: Станция управления | ||||
Встроенная антенна MPO |  36Gm (Мохо всегда. Ева большую часть времени. Дуна, если рядом) 36Gm (Мохо всегда. Ева большую часть времени. Дуна, если рядом) | |||
Расчет дальности
Отдельные суда не имеют фиксированной максимальной дальности: возможно ли соединение между двумя судами, находящимися на заданном расстоянии друг от друга, зависит от мощности передачи судов на оба конца соединения.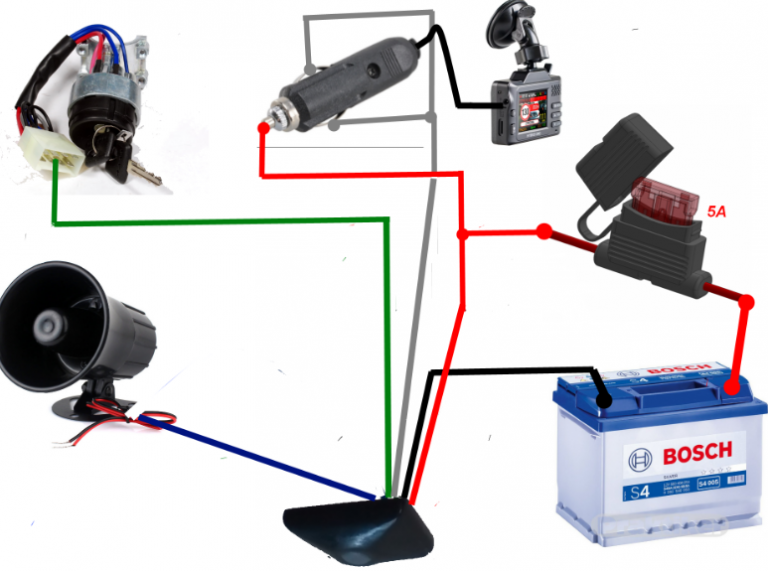 У каждой антенны есть «номинальная мощность», измеряемая безразмерным числом, которое влияет на общую номинальную мощность судна, к которому она прикреплена. Соединение может быть установлено между двумя судами, когда среднее геометрическое их номинальной мощности больше, чем их расстояние, измеренное в метрах.
У каждой антенны есть «номинальная мощность», измеряемая безразмерным числом, которое влияет на общую номинальную мощность судна, к которому она прикреплена. Соединение может быть установлено между двумя судами, когда среднее геометрическое их номинальной мощности больше, чем их расстояние, измеренное в метрах.
Это рассчитывается следующим образом:
Диапазон = Vessel1 × Vessel2 {\ displaystyle Range = {\ sqrt {Vessel_ {1} \ times Vessel_ {2}}}}
Где:
- Дальность {\ displaystyle Range} — это максимальное расстояние в метрах, на котором два судна могут соединиться друг с другом.
- Vessel1{\displaystyle Vessel_{1}} и Vessel2{\displaystyle Vessel_{2}} — номинальная мощность судов, пытающихся соединиться друг с другом.
Номинальная мощность антенн варьируется от 5k для встроенной антенны, доступной в каждом командном модуле, до 100G для самых больших тарелок. Сеть глубокого космоса Кербина можно рассматривать как судно с рейтингом 9.0003 2G / 50G / 250G (в зависимости от уровня станции слежения) для целей расчета дальности.
Сеть глубокого космоса Кербина можно рассматривать как судно с рейтингом 9.0003 2G / 50G / 250G (в зависимости от уровня станции слежения) для целей расчета дальности.
Если (в соответствии с настройками сложности) имеется только одна станция слежения, корабль должен зависеть от ретранслятора CommNet, установленного вручную вокруг Кербина, вот общие расстояния между антеннами.
| 5к | 36К† | 500 тыс. | 5М | 2G | 10G | 15G | 50G‡ | 100 г | 250 г‡ | |
|---|---|---|---|---|---|---|---|---|---|---|
| 5к | 5км | |||||||||
| 36к† | 13,4 км | 36 км | ||||||||
| 500к | 50км | 134. 2км 2км | 500км | |||||||
| 5М | 158.1км | 424,3 км | 1,6 мм | 5мм | ||||||
| 2G | 3,2 мм | 8,5 мм | 31,6 мм | 100мм | 2Gm | |||||
| 10G | 7,1 мм | 19мм | 70,7 мм | 223,6 мм | 4,5 Гм | 10 Гм | ||||
| 15G | 8,7 мм | 23,2 мм | 86,6 мм | 273,9 мм | 5,5 Гм | 12,2 Гм | 15 гм | |||
| 50G‡ | 15,8 мм | 42,4 мм | 158,1 мм | 500мм | 10 Гм | 22,4 Гм | 27,4 Гм | 50Gm | ||
| 100 г | 22,4 мм | 60мм | 223,6 мм | 707,1 мм | 14,1 Гм | 31,6 Гм | 38,7 Гм | 70,7 Гм | 100 гм | |
| 250G‡ | 35,4 мм | 94,9 мм | 353,6 мм | 1,1 Гм | 22,4 Гм | 50Gm | 61,2 Гм | 111,8 Гм | 158,1 Гм | 250 гм |
† Антенна 36K — это дополнительные наземные станции, которые могут быть обновлены или не обновлены с помощью KSC DSN. См. значения настроек сложности ниже.
См. значения настроек сложности ниже.
‡Антенны 50G и 250G являются частью KSC DSN, обновленной по сравнению со станцией слежения.
9{\text{Средний показатель сочетаемости}}}
Средний показатель сочетаемости судна – это среднее значение показателя сочетаемости всех антенн на судне, где вес каждой антенны пропорционален ее соответствующей номинальной мощности. Обратите внимание, что все антенны и реле имеют показатель степени сочетаемости 0,75, за исключением Communotron 16 и Communotron 16S со значениями 1,0 и 0,0 соответственно. Это означает, что средний показатель сочетаемости всегда будет равен 0,75 для судна, не содержащего ни одной из этих антенн. Для судов с Communotron 16 или 16S рассчитывается средний показатель сочетаемости:
Средний показатель сочетаемости = P1C1 + P2C2 + ⋯ + PnCnP1 + P2 + ⋯ + Pn {\ displaystyle {\ text {Средний показатель сочетаемости}} = {\ frac {P_ {1} C_ {1} + P_ {2} C_ {2 }+\cdots +P_{n}C_{n}}{P_{1}+P_{2}+\cdots +P_{n}}}}
Где:
- {P1,P2,…,Pn}{\displaystyle \{P_{1},P_{2},\dots ,P_{n}\}} — номинальная мощность каждой антенны.
- {C1,C2,…,Cn}{\displaystyle \{C_{1},C_{2},\dots ,C_{n}\}} — значения комбинируемости каждой соответствующей антенны.
Например, судно с Communotron 88-88 (мощность 100G, возможность комбинирования 0,75), а также Communotron 16 (мощность 500k, возможность комбинирования 1,00) будет иметь следующий средний показатель сочетаемости:
9{3})}}=0,75000125}
Значения показателя степени сочетаемости антенн можно найти в разделе «antennaCombinableExponent» в файлах конфигурации детали. Если значение отсутствует, используется значение по умолчанию 0,75.
Сила сигнала
Длина линии связи относительно ее максимальной дальности определяется силой сигнала. Уровень сигнала для соединения с одним переходом начинается со 100 % на нулевом расстоянии и уменьшается по мере увеличения расстояния до уровня 0 % на максимальном расстоянии соединения. Связь нелинейная: истинное значение требует нанесения относительного расстояния на кривой Безье с помощью клавиш «0 0 0 0» и «1 1 0 0». Это можно рассчитать следующим образом:
Это можно рассчитать следующим образом:
9{2}}
Результатом является мощность сигнала канала на указанном расстоянии. Обратите внимание, что вышеизложенное относится к каналу с одним переходом: если сигнал проходит через одно или несколько реле, уровень сигнала является произведением уровня сигнала каждого канала, составляющего путь к контрольной точке.
Пример
Расчет мощности сигнала между центром отслеживания DSN уровня 3 и зондом вокруг Джула на расстоянии 73,4 Гм от Кербина с двумя антеннами Communotron 88-88:
9{2}=0,707=71\%}
Таким образом, уровень сигнала для этого соединения будет 71%.
Передача науки
Если в настройках сложности включен CommNet, то передача науки через соединение с высоким уровнем сигнала даст бонус к эффективности передачи до 40% (до максимального значения 100%). Этот бонус уменьшается нелинейно с силой сигнала. Например, Гравитационное сканирование с низкой орбиты Джула даст 90*0,4=35 Science при передаче с отключенным CommNet или через маргинальное соединение, но при передаче через соединение с общей мощностью 100% даст 90*0,4*1,4=50,4 Наука.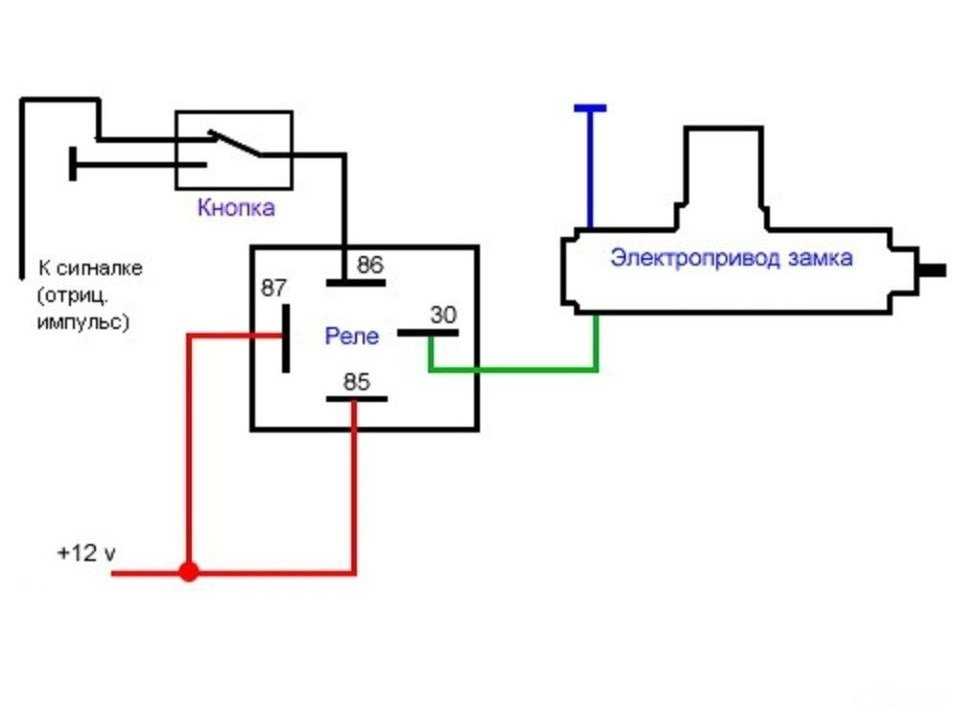
Соединения через ретрансляторы
Вы можете использовать корабли с ретрансляционными антеннами для усиления слабого сигнала или для передачи сигнала, когда ему препятствует небесное тело. Вам нужен только корабль с ядром зонда, антенной-ретранслятором, электричеством и способами его получения. Вам не нужно фокусироваться на корабле-ретрансляторе, чтобы передать сигнал, он будет передан автоматически.
При рассмотрении мощности антенны вашего судна, прямые антенны и антенны-ретрансляторы объединяются, чтобы дать вам «Мощность» для расчета мощности обратного сигнала к KSC, однако, если другое судно подключается через вас для связи с KSC, прямая Антенны не учитываются при любых расчетах.
Пример
- Судно с прямой антенной Communotron 88-88 и ретрансляционной антенной RA-100, вращающееся вокруг Eeloo на расстоянии 105Gm от KSC, с DSN уровня 3, соединится обратно с KSC с сигналом мощность 48%, так как обе антенны объединяются, чтобы дать повышенную мощность антенны для собственного прямого соединения.

- Судно, встретившееся с вышеупомянутым кораблем с жалким Коммунотроном 16, должно будет связаться с KSC через ретранслятор, поскольку его антенна недостаточно сильна, чтобы связаться с KSC напрямую. Поскольку этот небольшой зонд будет учитывать только антенну-ретранслятор RA-100 на корабле-ретрансляторе при расчете обратного соединения с KSC, номинальная мощность корабля-ретранслятора падает, и, следовательно, уровень сигнала упадет до 26% (обратите внимание на вычисления для этих чисел были скрыты от примера для облегчения понимания концепции).
Передача данных не потребляет электроэнергии, в отличие от передачи научных данных непосредственно с корабля, содержащего научные данные.
Игра всегда предпочитает прямое соединение с Кербином, даже если поблизости находится более мощное реле.
Ретрансляционные сети
Создание сети спутников вокруг каждой планеты может обеспечить непрерывную связь с KSC даже для кораблей, находящихся на дальней стороне тела.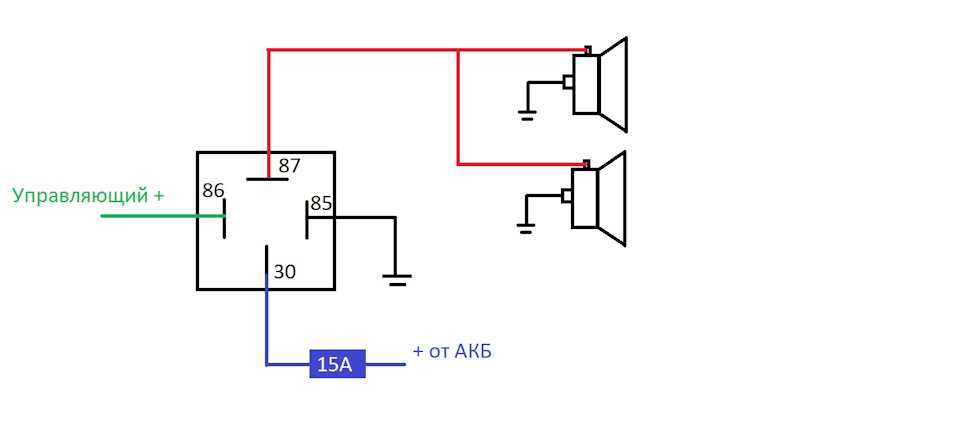 Для обеспечения непрерывного покрытия необходимо минимум три спутника-ретранслятора на тело. Спутники должны быть развернуты в форме равностороннего треугольника вдоль круговой орбиты, иначе могут быть периоды отключения электроэнергии. Минимальная высота для каждого спутника равна радиусу родительской планеты плюс высота самого высокого пика на поверхности. Самый простой способ создать такую сеть — построить эллиптическую орбиту для конвейера, где апоцентр — это желаемая высота круговой орбиты спутников, а перицентр — такой, чтобы общий период составлял 2/3 периода ретрансляции. Таким образом, каждый раз, когда конвейер достигает апоцентра, он может развернуть спутник-ретранслятор. Пример: Чтобы развернуть три спутника вокруг Кербина, эллиптическая орбита с высотами (2 868 751 м, 1 225 553 м) имеет период 4 часа, что составляет ровно 2/3 от 6 часов, что является периодом круговой орбиты с высота 2 868 751м.
Для обеспечения непрерывного покрытия необходимо минимум три спутника-ретранслятора на тело. Спутники должны быть развернуты в форме равностороннего треугольника вдоль круговой орбиты, иначе могут быть периоды отключения электроэнергии. Минимальная высота для каждого спутника равна радиусу родительской планеты плюс высота самого высокого пика на поверхности. Самый простой способ создать такую сеть — построить эллиптическую орбиту для конвейера, где апоцентр — это желаемая высота круговой орбиты спутников, а перицентр — такой, чтобы общий период составлял 2/3 периода ретрансляции. Таким образом, каждый раз, когда конвейер достигает апоцентра, он может развернуть спутник-ретранслятор. Пример: Чтобы развернуть три спутника вокруг Кербина, эллиптическая орбита с высотами (2 868 751 м, 1 225 553 м) имеет период 4 часа, что составляет ровно 2/3 от 6 часов, что является периодом круговой орбиты с высота 2 868 751м.
Главным преимуществом создания этих ретрансляционных сетей вокруг планет является то, что миссиям больше не нужно нести на каждой миссии мощную антенну; во многих ситуациях кораблю нужен только Communotron 16 или даже встроенный 5K, и он может полагаться на реле при входе в SOI цели.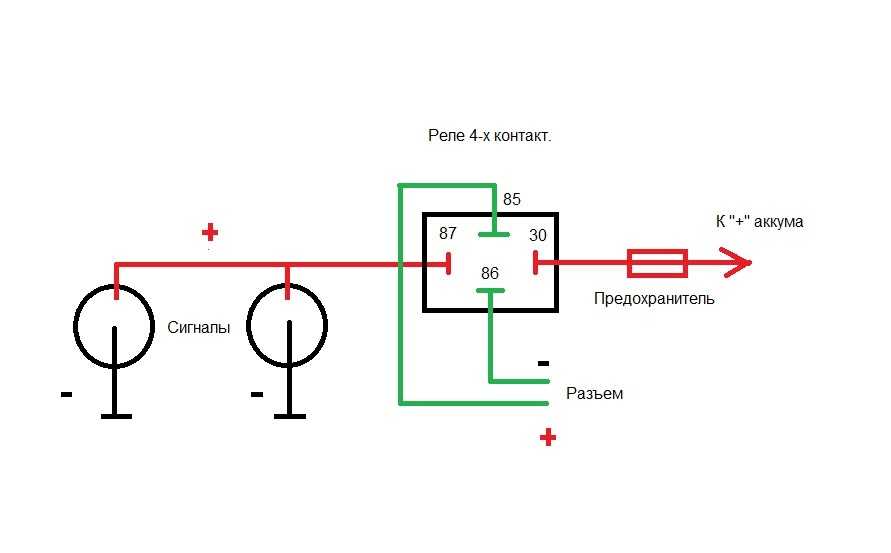 Все луны могут достичь своей родительской планеты только с помощью антенны с высоким коэффициентом усиления HG-5, при условии, что на орбите вокруг родительской планеты есть ретрансляционная сеть, поэтому нужно беспокоиться только о расчете связи между планетой и KSC. Пример: HG-5 на орбите Pol достаточно силен, чтобы соединиться с RA-15 на орбите Jool, что означает, что корабль, работающий в SOI Pol, может полагаться на встроенную антенну 5K для поддержания связи с KSC через DSN, а не быть обремененным. с более мощной антенной.
Все луны могут достичь своей родительской планеты только с помощью антенны с высоким коэффициентом усиления HG-5, при условии, что на орбите вокруг родительской планеты есть ретрансляционная сеть, поэтому нужно беспокоиться только о расчете связи между планетой и KSC. Пример: HG-5 на орбите Pol достаточно силен, чтобы соединиться с RA-15 на орбите Jool, что означает, что корабль, работающий в SOI Pol, может полагаться на встроенную антенну 5K для поддержания связи с KSC через DSN, а не быть обремененным. с более мощной антенной.
В следующей таблице указаны минимальная мощность антенны и конфигурации для каждой из планет.
| Планета | Максимальное расстояние от Кербина | Минимальная сила ретрансляции для канала DSN | ||
|---|---|---|---|---|
| 2G | 50 г | 250 г | ||
| Мохо | 18 862 978 560 м | 177 905 980 078📡 | 7 116 239 204📡 | 1 423 247 841📡 |
| (РА-100 @ 2) + (РА-15 @ 2) | РА-15 | РА-2 | ||
| Ева | 23 530 851 643м | 276 850 489 523📡 | 11 074 019 581📡 | 2 214 803 917📡 |
| РА-100 @ 4 | РА-15 | РА-2 @ 2 | ||
| Дуна | 35 383 029 419м | 625 979 385 433📡 | 25 039 175 418📡 | 5 007 835 084📡 |
| Невозможно | РА-15 @ 2 | РА-2 @ 4 | ||
| Платье | 60 360 893 643м | 1 821 718 740 691📡 | 72 868 749 628📡 | 14 573 749 926📡 |
| Невозможно | РА-100 | РА-15 | ||
| Джул | 85 812 078 643м | 3 681 856 420 517📡 | 147 274 256 821📡 | 29 454 851 365📡 |
| Невозможно | РА-100 @ 2 | РА-15 @ 3 | ||
| Илу | 127 149 553 456м | 8 083 504 472 031📡 | 323 340 178 882📡 | 64 668 035 777📡 |
| Невозможно | РА-100 @ 5 | РА-100 | ||
Последствия потери сигнала
В зависимости от настроек сложности при потере сигнала CommNet возникает несколько эффектов либо от KSC, либо от контрольной точки зонда.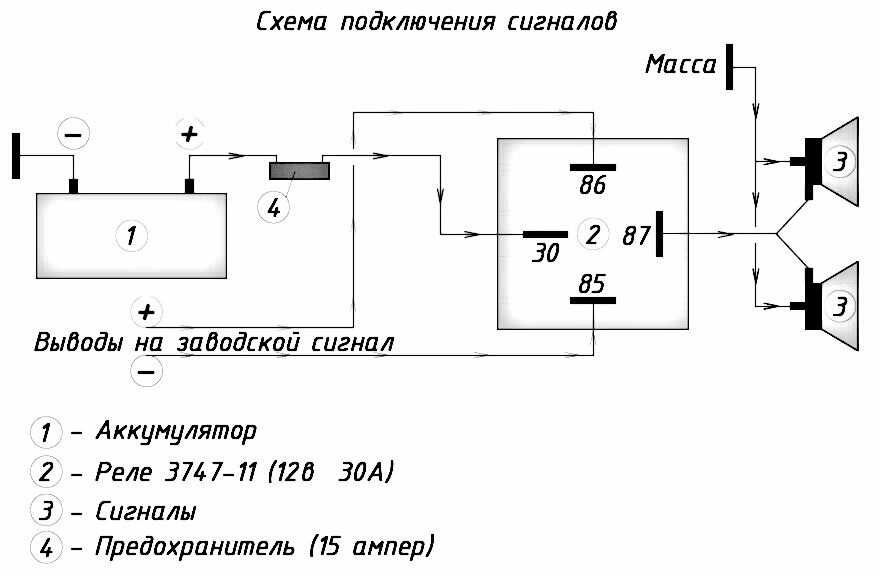 Это может быть связано либо с потерей электроэнергии, выходом за пределы диапазона CommNet, либо с разрушением антенны. Существует также настройка, в которой вы можете отключить сигнал при слишком быстром входе в атмосферу тела.
Это может быть связано либо с потерей электроэнергии, выходом за пределы диапазона CommNet, либо с разрушением антенны. Существует также настройка, в которой вы можете отключить сигнал при слишком быстром входе в атмосферу тела.
- Пилотируемый без пилота : вы потеряете возможность создавать узлы маневра или удалять существующие шаг), точное управление тягой (только 0 или максимум) и возможность создавать или удалять узлы маневра. Если ваш зонд способен зафиксировать ось маневра через SAS, рекомендуется создать узел маневра до отключения передачи (прохождение за телом), чтобы позволить кораблю выполнить маневр.
- Зонд, требуется управление проверено : Вы полностью потеряете контроль над зондом. Либо подождите, пока он вернется в радиус действия CommNet, либо отправьте корабль с контрольной точкой зонда для выполнения общих маневров (установка солнечных батарей, точное управление и т. д.).
SAS не передается KSC или контрольной точкой проверки. Если вы не можете использовать SAS, проверьте, является ли ваш датчик управляемым (батареи, диапазон CommNet) или есть ли на борту пилот. Уровень SAS зависит от самого корабля (пробные уровни SAS или уровень пилота).
Если вы не можете использовать SAS, проверьте, является ли ваш датчик управляемым (батареи, диапазон CommNet) или есть ли на борту пилот. Уровень SAS зависит от самого корабля (пробные уровни SAS или уровень пилота).
Значения настроек сложности
| Настройка | При ВЫКЛ. | При включении | Настройка на уровень сложности | |||
|---|---|---|---|---|---|---|
| Легко | Нормальный | Умеренный | Жесткий | |||
| Активировать CommNet | Все корабли имеют полный контроль и могут строить узлы маневрирования, где бы они ни находились. Если у них есть антенна, они могут передавать научные данные откуда угодно. | Корабли ведут себя по-разному в зависимости от статуса CommNet, см. ниже | Выкл. | Вкл. | Вкл. | Вкл. |
| Необходимый сигнал для управления | Когда корабли, предназначенные только для зондов, выходят за пределы досягаемости, они переходят в частичный контроль (так же, как и в режим гибернации), где они могут только привязываться к доступной оси SAS и дросселировать на 0 или 100%.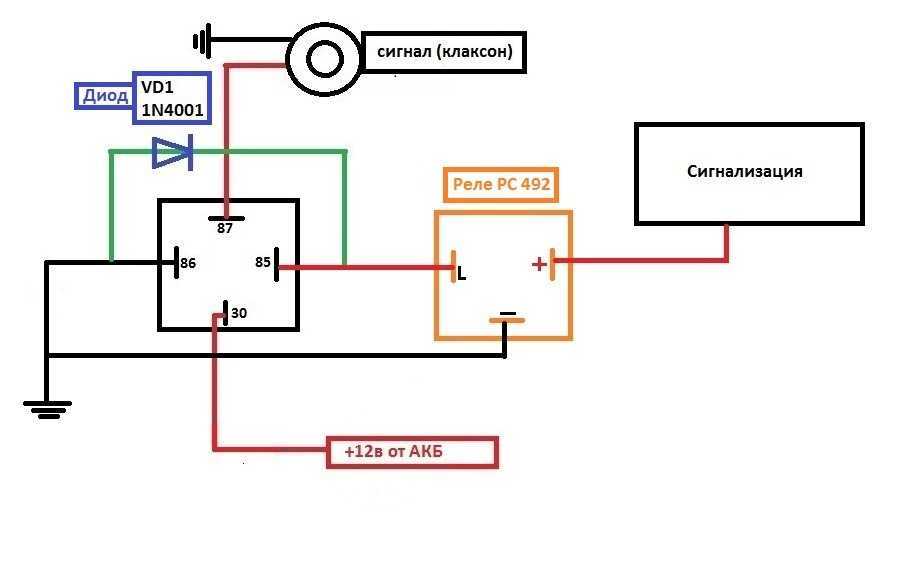 | Находясь вне досягаемости, корабли, предназначенные только для зондов, полностью теряют управление. | – | Выкл. | Выкл. | Выкл. |
| Плазменное затемнение | Повторный вход в атмосферу не влияет на CommNet | Повторный вход в атмосферу снижает уровень сигнала, вы можете навести значок сети на панели телеметрии, чтобы увидеть, сколько | – | Выкл. | Выкл. | Выкл. |
| Модификатор диапазона | Значение этого ползунка представляет собой значение множителя, которое применяется к уровням мощности всех антенн. | – | 1,00 | 0,80 | 0,65 | |
| Модификатор DSN | Значение этого ползунка представляет собой значение множителя, которое применяется к уровню мощности сети DSN. | – | 1,00 | 1,00 | 1,00 | |
| Модификатор окклюзии, вакуум | Значение этого ползунка представляет собой значение множителя, которое применяется к эффективному размеру безатмосферных тел, которые могут блокировать сигналы между антеннами. | – | 0,90 | 1,00 | 1,00 | |
| Модификатор окклюзии, атм. | Значение этого ползунка представляет собой значение множителя, которое применяется к эффективному размеру тел с атмосферой, которая может блокировать сигналы между антеннами. | – | 0,75 | 0,85 | 1,00 | |
| Включить дополнительные наземные станции | Только одна наземная станция имеет DSN: Космический центр Кербала. Другие наземные станции имеют рейтинг антенны по умолчанию ~ 36k *. | В Кербине есть много наземных станций, которые имеют тот же рейтинг DNS, что и Станция слежения. | – | Вкл. | Вкл. | Вкл. |
- Судя по косвенным данным, по умолчанию DSN отключен рейтинг антенны наземной станции составляет приблизительно 36k, но CommNet предпочитает дополнительные прыжки DSN на Кербине, если доступен путь прохождения сигнала.
Диапазоны CommNet для отключенных DSN наземных станций:
| 5к | 500 тыс. | 5М | 2G | 10G | 15G | 100 г | |
|---|---|---|---|---|---|---|---|
| 36к | 13.42км | 134.16км | 424,26 км | 8,49 мм | 18,97 мм | 23,24 мм | 60мм |
Учебные пособия, калькуляторы и примеры
Документ Google Sheets, который поможет вам рассчитать мощность сигнала и диапазоны, можно найти здесь: CommNet Signal Strength Calculator & Antenna Selector
Интерактивный инструмент для визуализации мощности сигнала в зависимости от положения планет можно найти здесь: KSP Visual Calculator
Примеры орбит для создания планетарной сети можно найти по адресу: Учебное пособие: Идеальные орбиты для спутников связи
Один из методов настройки полносистемной сети можно найти в учебнике: Настройка системы CommNet.
Сигнальное реле во время хемотаксиса — PMC
- Список журналов
- Рукописи авторов HHS
- PMC2745292
Дж Микроск. Авторская рукопись; доступно в PMC 2009 16 сентября.
Опубликовано в окончательной редакции как:
J Microsc. 2008 сен; 231(3): 529–534.
doi: 10.1111/j.1365-2818.2008.02066.x
PMCID: PMC2745292
NIHMSID: NIHMS126070
PMID: 18755009
Информация об авторе Информация об авторских правах и лицензии Отказ от ответственности
Способность клеток мигрировать в ответ на внешние сигналы, процесс, известный как хемотаксис. Он проявляется широким разнообразием типов клеток в контексте эмбриогенеза, ангиогенеза, воспаления, заживления ран и многих других сложных физиологических процессов.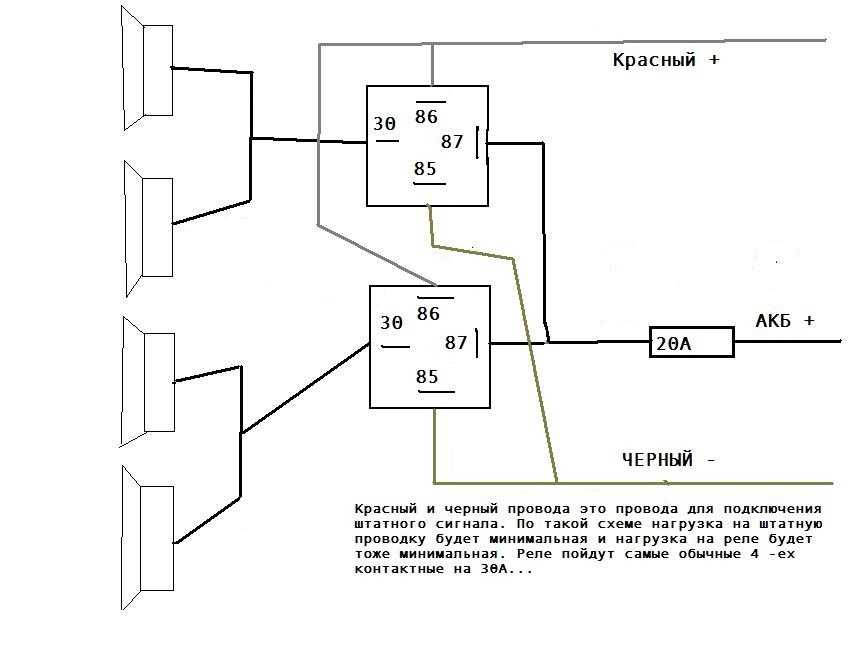 Здесь мы обсуждаем сигналы, которые контролируют направленную миграцию социальных амеб Dictyostelium discoideum как в виде одиночных клеток, так и в условиях групповой миграции. Этот многоклеточный организм послужил превосходной модельной системой для расшифровки амебоидоподобной миграции лейкоцитов и сыграл ключевую роль в установлении сигнальных парадигм в области хемотаксиса. Мы предвидим, что Dictyostelium будет продолжать продвигать базовые знания, поскольку мы стремимся понять механизмы, регулирующие миграцию групповых клеток.
Здесь мы обсуждаем сигналы, которые контролируют направленную миграцию социальных амеб Dictyostelium discoideum как в виде одиночных клеток, так и в условиях групповой миграции. Этот многоклеточный организм послужил превосходной модельной системой для расшифровки амебоидоподобной миграции лейкоцитов и сыграл ключевую роль в установлении сигнальных парадигм в области хемотаксиса. Мы предвидим, что Dictyostelium будет продолжать продвигать базовые знания, поскольку мы стремимся понять механизмы, регулирующие миграцию групповых клеток.
Ключевые слова: цАМФ, хемотаксис, Dictyostelium discoideum
D. discoideum — наиболее изученный вид социальных амеб, рано дивергировавший после расщепления растений и животных и представляющий собой один из базальных кластеров коронных групп эукариот (Kessin, 2001; Baldauf , 2003). Его небольшой геном (34 Mbp) был полностью секвенирован (Eichinger et al. , 2005). Он организован в виде шести хромосом, содержащих примерно 12 000 генов, многие из которых имеют гомологию с высшими эукариотами (Fey и др.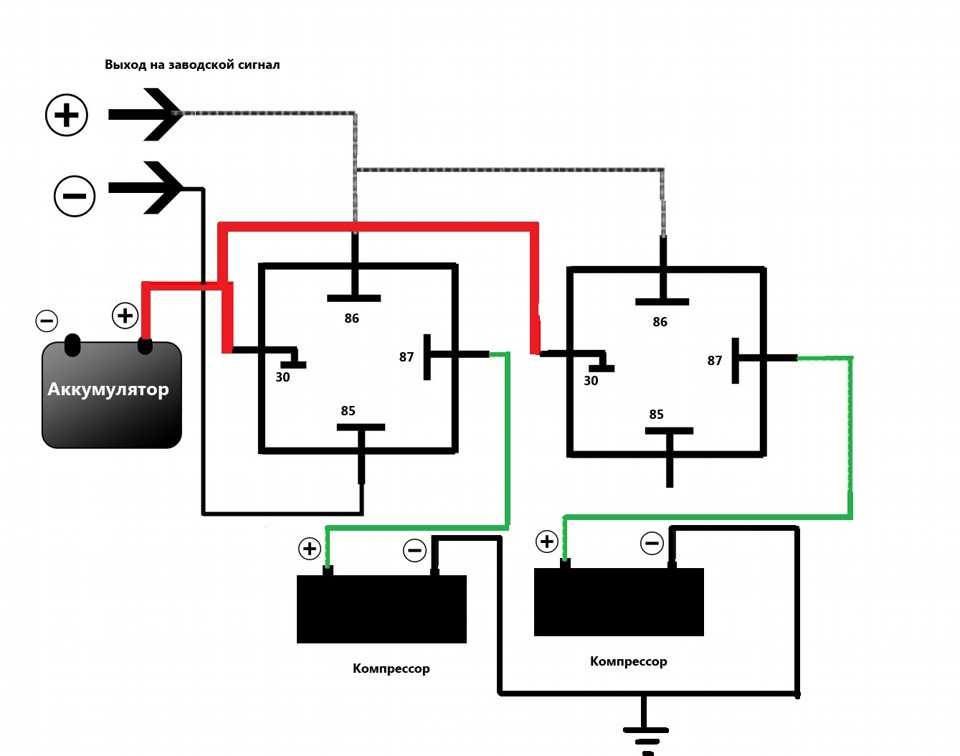 , 2006). Сила Dictyostelium как модельного организма заключается в его легкодоступной генетике и биохимии. Амебы можно выращивать во встряхиваемых культурах, и за несколько дней можно получить 10 12 клональных клеток для биохимических анализов. Несущественные гены могут быть разрушены путем гомологичной рекомбинации или подавлены с помощью РНКи, а методы инсерционного мутагенеза позволяют идентифицировать новые гены (Landree & Devreotes, 2004; Kimmel & Faix, 2006; Kuhlmann 9).1031 и др. , 2006; Куспа, 2006). Наконец, легко получить трансгенные клетки, экспрессирующие мутированные гены или слияния GFP, и они оказались бесценными для расшифровки функции белка и пространственно-временного распределения белков в живых клетках (Muller-Taubenberger, 2006). Генетическая и литературная база данных о Dictyostelium , Dictybase (http://dictybase.org) и центральное хранилище штаммов и плазмид, Dictyostelium Stock Center, делает Dictyostelium исследования очень доступны (Chisholm et al.
, 2006). Сила Dictyostelium как модельного организма заключается в его легкодоступной генетике и биохимии. Амебы можно выращивать во встряхиваемых культурах, и за несколько дней можно получить 10 12 клональных клеток для биохимических анализов. Несущественные гены могут быть разрушены путем гомологичной рекомбинации или подавлены с помощью РНКи, а методы инсерционного мутагенеза позволяют идентифицировать новые гены (Landree & Devreotes, 2004; Kimmel & Faix, 2006; Kuhlmann 9).1031 и др. , 2006; Куспа, 2006). Наконец, легко получить трансгенные клетки, экспрессирующие мутированные гены или слияния GFP, и они оказались бесценными для расшифровки функции белка и пространственно-временного распределения белков в живых клетках (Muller-Taubenberger, 2006). Генетическая и литературная база данных о Dictyostelium , Dictybase (http://dictybase.org) и центральное хранилище штаммов и плазмид, Dictyostelium Stock Center, делает Dictyostelium исследования очень доступны (Chisholm et al.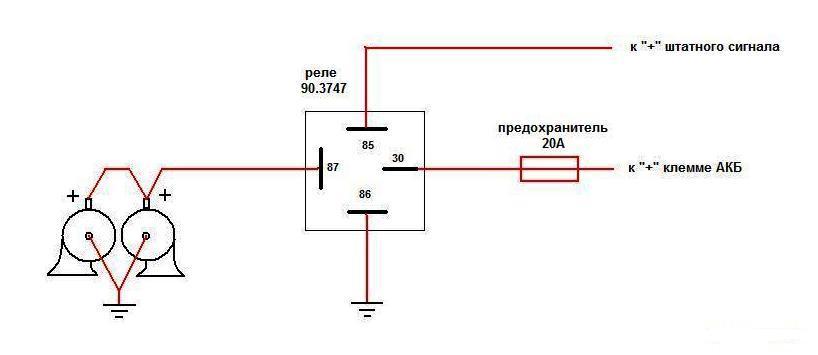 , 2006).
, 2006).
Клетки Dictyostelium живут в двух независимых состояниях. При наличии питательных веществ они ведут одиночный образ жизни, развиваясь автономно. Однако при голодании клетки перестают делиться и вступают в программу развития, в которой до 100 000 клеток объединяются и дифференцируются в многоклеточный организм — плодовое тело, состоящее из спор на стебле вакуолизированных клеток (). Возвращение благоприятных питательных условий приводит к прорастанию спор и повторному запуску цикла (Williams & Harwood, 2003; Chisholm & Firtel, 2004; Kimmel & Firtel, 2004). Хемотаксис необходим на протяжении всего жизненного цикла Диктиостелиум . В своей фазе роста клетки охотятся и фагоцитируют бактерии (их любимый источник пищи), обнаруживая и мигрируя к фолиевой кислоте, побочному продукту бактериального метаболизма. На раннем этапе развития цАМФ становится основным хемоаттрактантом, и клетки используют хемотаксис, чтобы собираться вместе и образовывать агрегаты.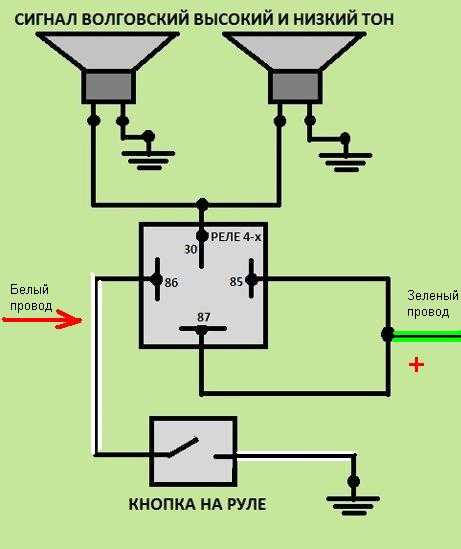 Интересно, что когда клетки мигрируют, они быстро организуются в группы, в которых отдельные клетки выстраиваются в линию голова к хвосту, образуя цепочки клеток или потоки, которые в конечном итоге образуют агрегаты (Mahadeo & Parent, 2006) (4). На более поздних стадиях развития клетки продолжают группами реагировать на внешние сигналы цАМФ, вращаясь и демонстрируя периодическую поляризацию и всплески движения (Weijer, 2004). С момента открытия Dictyostelium предоставил очень полезную и, возможно, беспрецедентную модель для изучения подвижности и хемотаксиса.
Интересно, что когда клетки мигрируют, они быстро организуются в группы, в которых отдельные клетки выстраиваются в линию голова к хвосту, образуя цепочки клеток или потоки, которые в конечном итоге образуют агрегаты (Mahadeo & Parent, 2006) (4). На более поздних стадиях развития клетки продолжают группами реагировать на внешние сигналы цАМФ, вращаясь и демонстрируя периодическую поляризацию и всплески движения (Weijer, 2004). С момента открытия Dictyostelium предоставил очень полезную и, возможно, беспрецедентную модель для изучения подвижности и хемотаксиса.
Открыть в отдельном окне
Жизненный цикл социальной амебы Dictyostelium . (A) Монтаж изображений, изображающих программу развития голодающих клеток Dictyostelium . Клетки высевали на чашки с непитательным агаром и делали снимки в указанное время после начала голодания. Бар представляет собой 1 мм. (B) Изображения, показывающие клетки, мигрирующие потоками во время хемотаксиса. На изображении слева изображено несколько центров агрегации в потоках.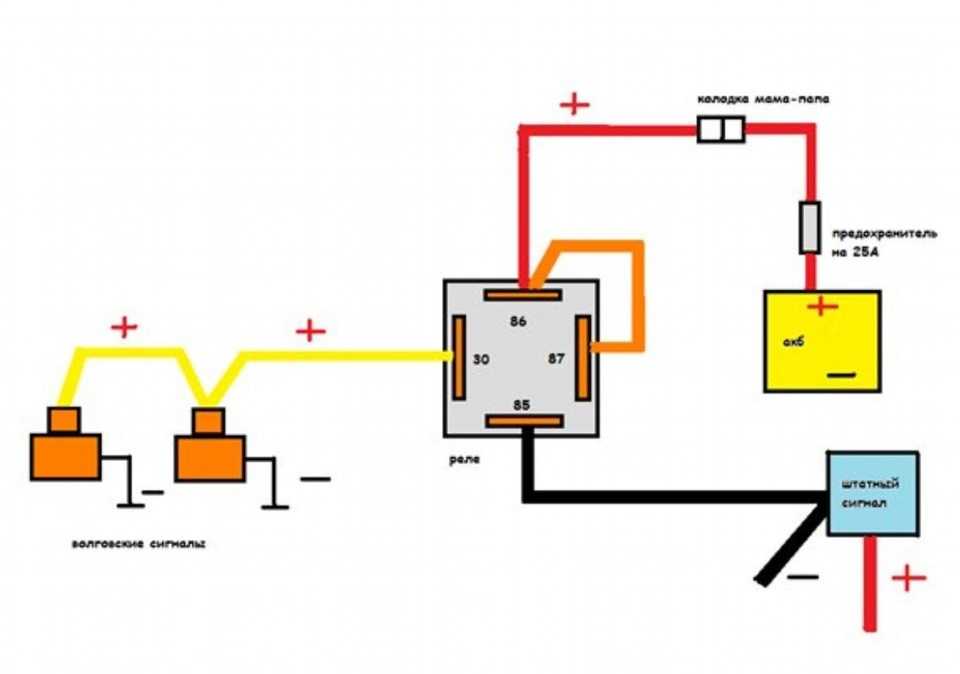 На рисунке справа представлено увеличенное изображение клеток, мигрирующих потоками в микропипетку, содержащую 10 мкМ цАМФ. Белая стрелка указывает положение микропипетки. Полоса соответствует 1 мм для правого изображения и 250 мкм для левого изображения.
На рисунке справа представлено увеличенное изображение клеток, мигрирующих потоками в микропипетку, содержащую 10 мкМ цАМФ. Белая стрелка указывает положение микропипетки. Полоса соответствует 1 мм для правого изображения и 250 мкм для левого изображения.
Пути, сохранившиеся на протяжении эволюции, передают сигналы, регулирующие весь жизненный цикл Dictyostelium . При голодании четыре различных, но очень родственных рецептора цАМФ (cAR1-4) по-разному экспрессируются, чтобы опосредовать эффекты цАМФ (Hereld & Devreotes, 1992; Kim et al. , 1998). Эти рецепторы являются частью большого семейства из семи трансмембранных рецепторных белков, которые передают свои эффекты через гетеротримерные G-белки — рецепторы, связанные с G-белками (GPCR) (Karnik 9).1031 и др. , 2003; Миллиган и Костенис, 2006 г .; Прабху и Эйхингер, 2006). В клетках млекопитающих большое семейство хемокинов опосредует хемотаксические ответы. Хемокины представляют собой небольшие пептиды, состоящие примерно из 100 аминокислот. Они классифицируются как CXC или CC в соответствии с положением первых двух остатков цистеина в последовательности хемокинов (Baggiolini, 2001). Подобно цАМФ в Dictyostelium , хемокины опосредуют свою хемотаксическую активность через GPCR, где связывание хемоаттрактанта с их специфическим рецептором приводит к диссоциации G-белков на α — и βγ -субъединиц, а также на активацию различных консервативных эффекторов (Bagorda et al. , 2006).
Они классифицируются как CXC или CC в соответствии с положением первых двух остатков цистеина в последовательности хемокинов (Baggiolini, 2001). Подобно цАМФ в Dictyostelium , хемокины опосредуют свою хемотаксическую активность через GPCR, где связывание хемоаттрактанта с их специфическим рецептором приводит к диссоциации G-белков на α — и βγ -субъединиц, а также на активацию различных консервативных эффекторов (Bagorda et al. , 2006).
В Dictyostelium межклеточная коммуникация необходима, чтобы позволить клеткам спонтанно мигрировать в определенном направлении и образовывать агрегаты. С этой целью организм разработал впечатляющий механизм для тонкой регуляции обнаружения, синтеза, секреции и деградации цАМФ. Три различные аденилатциклазы экспрессируются на протяжении всего жизненного цикла Dictyostelium , ACA, ACB и ACG (Saran et al. , 2002; Kriebel & Parent, 2004). Во время ранней агрегации, когда клетки проявляют самую сильную хемотаксическую активность, АКА максимально экспрессируется.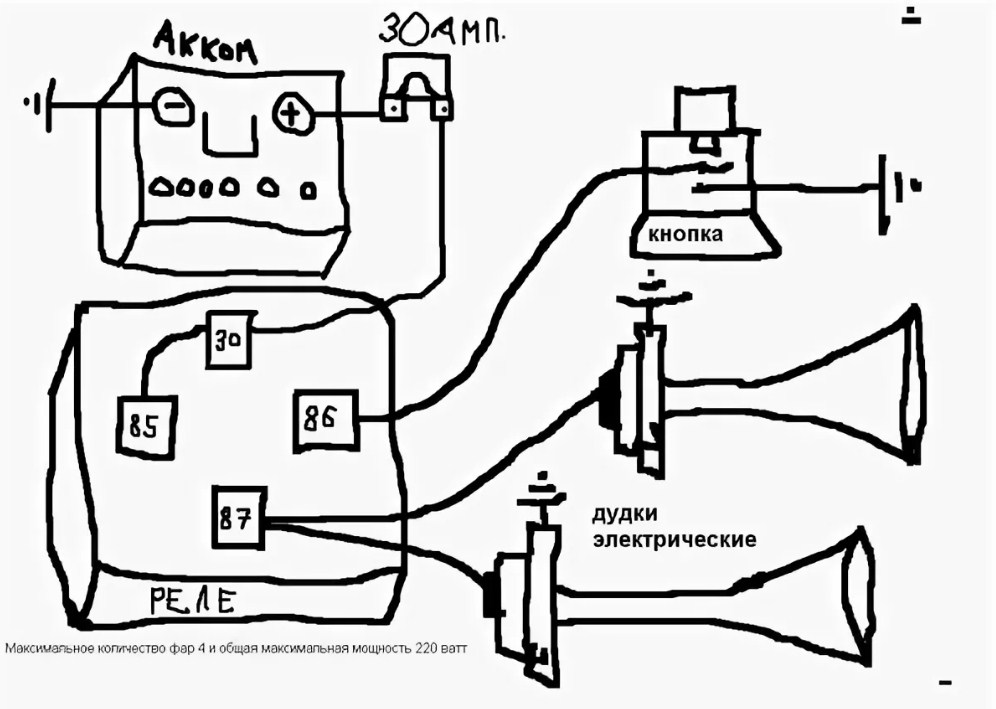 ACA родственна аденилатциклазам, связанным с G-белком млекопитающих, и состоит из двух наборов из шести трансмембранных доменов, за каждым из которых следует большая цитозольная петля, в которой находятся каталитические домены (4). Связывание цАМФ с цАР1 приводит к активации многих эффекторов, в том числе АСА, которая превращает АТФ в цАМФ. Хотя часть продуцируемого цАМФ остается внутри клетки для активации нижестоящих путей, большая часть цАМФ фактически секретируется для привлечения соседних клеток (Dinauer 9).1031 и др. , 1980а; Махадео и Родитель, 2006) (). Это включает в себя петлю ретрансляции сигнала, которая необходима для агрегации клеток. Действительно, клетки, в которых отсутствует какой-либо из компонентов, ведущих к выработке цАМФ, не могут начать развиваться и остаются в виде гладких монослоев клеток при высеве на непитательные поверхности (Pitt et al. , 1992; Insall et al. , 1994; Chen и др. , 1997).
ACA родственна аденилатциклазам, связанным с G-белком млекопитающих, и состоит из двух наборов из шести трансмембранных доменов, за каждым из которых следует большая цитозольная петля, в которой находятся каталитические домены (4). Связывание цАМФ с цАР1 приводит к активации многих эффекторов, в том числе АСА, которая превращает АТФ в цАМФ. Хотя часть продуцируемого цАМФ остается внутри клетки для активации нижестоящих путей, большая часть цАМФ фактически секретируется для привлечения соседних клеток (Dinauer 9).1031 и др. , 1980а; Махадео и Родитель, 2006) (). Это включает в себя петлю ретрансляции сигнала, которая необходима для агрегации клеток. Действительно, клетки, в которых отсутствует какой-либо из компонентов, ведущих к выработке цАМФ, не могут начать развиваться и остаются в виде гладких монослоев клеток при высеве на непитательные поверхности (Pitt et al. , 1992; Insall et al. , 1994; Chen и др. , 1997).
Открыть в отдельном окне
Путь передачи сигнала цАМФ в Диктиостелиум .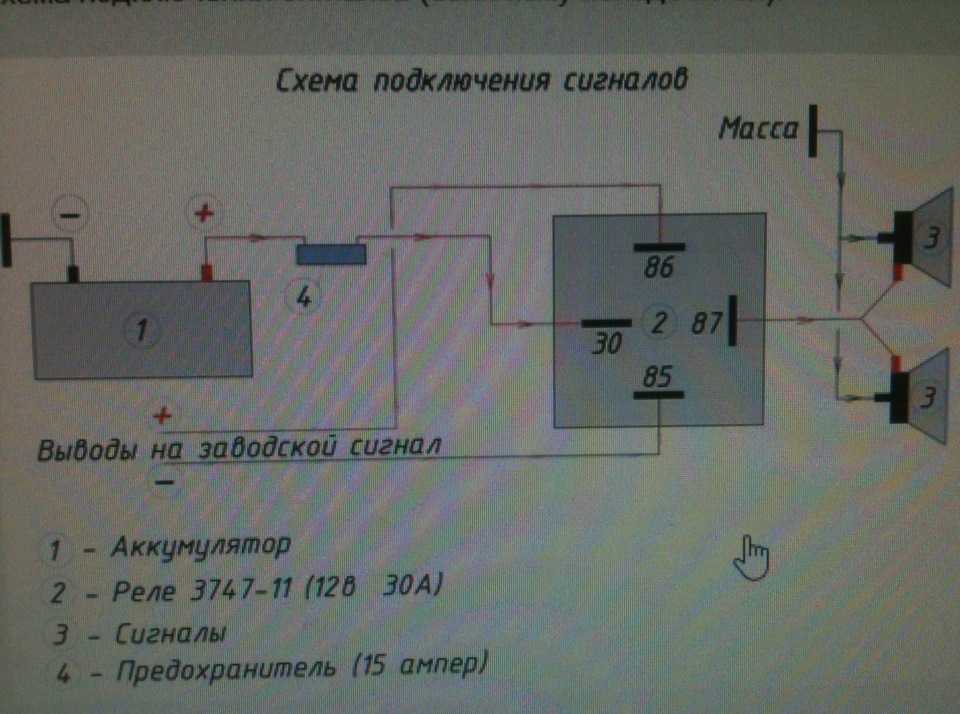 (A) Мультфильм, представляющий каскад событий, ведущих к активации ВДА. (B) Мультфильм, изображающий клеточное распределение различных компонентов сигнальной трансдукции во время хемотаксиса. Фактические изображения клеток, экспрессирующих CRAC-GFP или ACA-YFP, представлены на последних двух панелях. Подробности смотрите в тексте.
(A) Мультфильм, представляющий каскад событий, ведущих к активации ВДА. (B) Мультфильм, изображающий клеточное распределение различных компонентов сигнальной трансдукции во время хемотаксиса. Фактические изображения клеток, экспрессирующих CRAC-GFP или ACA-YFP, представлены на последних двух панелях. Подробности смотрите в тексте.
Два различных механизма позволяют клеткам сохранять чувствительность к цАМФ. Во-первых, опосредованная хемоаттрактантом активация АЦА носит временный характер, демонстрируя начальный пик активации, за которым следует фаза адаптации, приводящая к снижению количества продуцируемого цАМФ (Dinauer 9).1031 и др. , 1980b). Механизмы, регулирующие этот важный адаптационный ответ, еще предстоит выяснить. Было показано, что адаптация ACA не зависит от фосфорилирования cARs и, по-видимому, происходит после G-белков (Lilly & Devreotes, 1995; Parent & Devreotes, 1996). Во-вторых, после образования цАМФ легко расщепляется различными внутриклеточными и внеклеточными фосфодиэстеразами (Saran et al. , 2002; Bader et al. , 2007) (). Это позволяет системе вернуться к базальным уровням и отреагировать на новый цикл активации.
, 2002; Bader et al. , 2007) (). Это позволяет системе вернуться к базальным уровням и отреагировать на новый цикл активации.
Механизмы, которые регулируют опосредованную хемоаттрактантом активацию АЦА, сложны (). Каскад инициируется на плазматической мембране, где G βγ -субъединицы активируют PI3K Ras-зависимым образом (Sasaki et al. , 2004). Это приводит к образованию фосфатидилинозитол-3,4,5-фосфата (PIP 3 ) и привлечению белка CRAC, содержащего домен гомологии плекстрина (PH) (цитозольный регулятор аденилатциклазы) (Parent et al. 9).1032, 1998). Первоначально CRAC был выделен в результате скрининга химического мутагенеза, и было показано, что он необходим для опосредованной хемоаттрактантом активации ACA (Theibert & Devreotes, 1986; Insall et al. , 1994; Lilly & Devreotes, 1994). Совсем недавно мы показали, что CRAC независимо регулирует хемотаксис и активацию ACA ниже PI3K (Comer et al. , 2005). Механизмы, с помощью которых CRAC активирует ACA и хемотаксис, еще предстоит определить.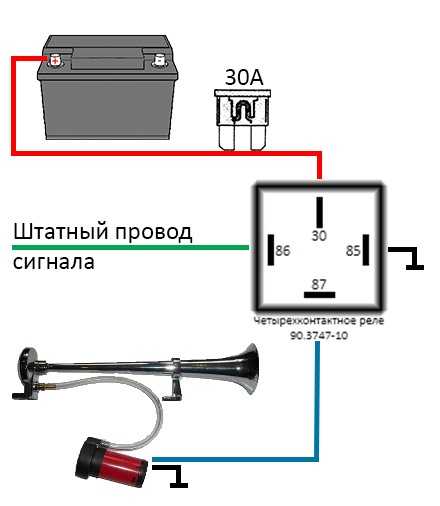 Однако промежуточные компоненты, скорее всего, участвуют в контроле активности АЦА, поскольку опосредованная хемоаттрактантом транслокация CRAC на плазматическую мембрану происходит за полную 1 мин до пика активности АЦА (родительский элемент 9).1031 и др. , 1998).
Однако промежуточные компоненты, скорее всего, участвуют в контроле активности АЦА, поскольку опосредованная хемоаттрактантом транслокация CRAC на плазматическую мембрану происходит за полную 1 мин до пика активности АЦА (родительский элемент 9).1031 и др. , 1998).
Мишень рапамицинового комплекса 2 (TORC2) также является ключевым регулятором хемотаксиса и активности ACA у Dictyostelium (). TOR является членом семейства фосфатидилкиназ серин-треониновых протеинкиназ. Он существует в двух функционально различных белковых комплексах (TORC1 и TORC2) (Sarbassov et al. , 2005; Wullschleger et al. , 2006; Yang & Guan, 2007). В клетках млекопитающих TORC1, чувствительный к рапамицину, состоит из Raptor, LST8, PRAS40 и TOR и регулирует синтез белка и рост клеток. TORC2 нечувствителен к рапамицину. Он состоит из Rictor, LST8, Sin1 и TOR и в основном участвует в регуляции организации цитоскелета. В Dictyostelium экспрессируются гомологи TOR, Raptor, LST8, Rictor (называемый Pianissimo) и Sin1 (называемый Rip3), и особенно хорошо охарактеризована роль TORC2 в хемотаксисе и передаче сигнала (Chen et al. , 1997; Lee и др. , 1999; Lee и др. , 2005; Sasaki & Firtel, 2006). Клетки, лишенные Pianissimo (Pia), LST8 или Rip3, имеют общий фенотип, демонстрирующий сильные дефекты в опосредованной хемоаттрактантом активации ACA, а также в хемотаксисе. Эти дефекты зависят от наличия предварительно сформированного TORC2. Используя эксперименты по восстановлению, было показано, что CRAC не требуется для образования TORC2 (Lee и др. , 2005). Тем не менее, эксперименты, проведенные на клетках, лишенных как Pia, так и CRAC, установили, что активность ACA абсолютно требует участия обоих путей (Chen et al. , 1997). Интересно, что работа, первоначально выполненная на Dictyostelium , показала, что TORC2 также необходим для опосредованной хемоаттрактантом активации Akt/PKB и родственного PKBR1 (Lee et al. , 2005). Хотя было показано, что клетки, лишенные Akt/PKB и PKBR1, имеют дефекты хемотаксиса и полярности, не было определено, регулируют ли эти эффекторы также ACA (Meili 9).
, 1997; Lee и др. , 1999; Lee и др. , 2005; Sasaki & Firtel, 2006). Клетки, лишенные Pianissimo (Pia), LST8 или Rip3, имеют общий фенотип, демонстрирующий сильные дефекты в опосредованной хемоаттрактантом активации ACA, а также в хемотаксисе. Эти дефекты зависят от наличия предварительно сформированного TORC2. Используя эксперименты по восстановлению, было показано, что CRAC не требуется для образования TORC2 (Lee и др. , 2005). Тем не менее, эксперименты, проведенные на клетках, лишенных как Pia, так и CRAC, установили, что активность ACA абсолютно требует участия обоих путей (Chen et al. , 1997). Интересно, что работа, первоначально выполненная на Dictyostelium , показала, что TORC2 также необходим для опосредованной хемоаттрактантом активации Akt/PKB и родственного PKBR1 (Lee et al. , 2005). Хотя было показано, что клетки, лишенные Akt/PKB и PKBR1, имеют дефекты хемотаксиса и полярности, не было определено, регулируют ли эти эффекторы также ACA (Meili 9).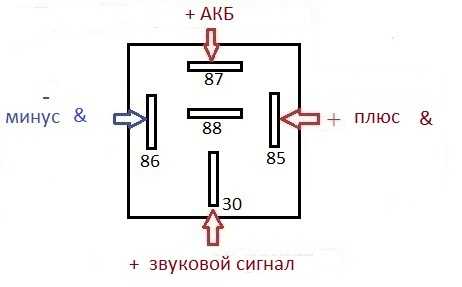 1031 и др. , 2000).
1031 и др. , 2000).
Благодаря использованию технологии GFP стало возможным визуализировать распределение большинства сигнальных компонентов цАМФ в живых хемотаксирующих клетках (). Хотя и рецепторы, и G-белки остаются равномерно распределенными на плазматической мембране во время этого процесса, обнаружено, что Ras, PI3K, PKB и CRAC специфически и динамично перемещаются из цитозоля к переднему краю хемотаксирующих клеток (Xiao et al. , 1997). ; родитель и др. , 1998; Мейли и др. , 1999; Джин и др. , 2000; Фунамото и др. , 2002; Сасаки и др. , 2004) Одновременно фосфатаза, ответственная за дефосфорилирование PIP 3, PTEN, также динамически связана с плазматической мембраной по бокам и сзади клеток (Iijima & Devreotes, 2002). Это строго регулируемое и комплементарное клеточное распределение PI3K и PTEN делает возможной ограниченную продукцию PIP 3 на переднем крае клеток, где перераспределяются CRAC и другие белки, содержащие домен PH (Comer&Parent, 2002).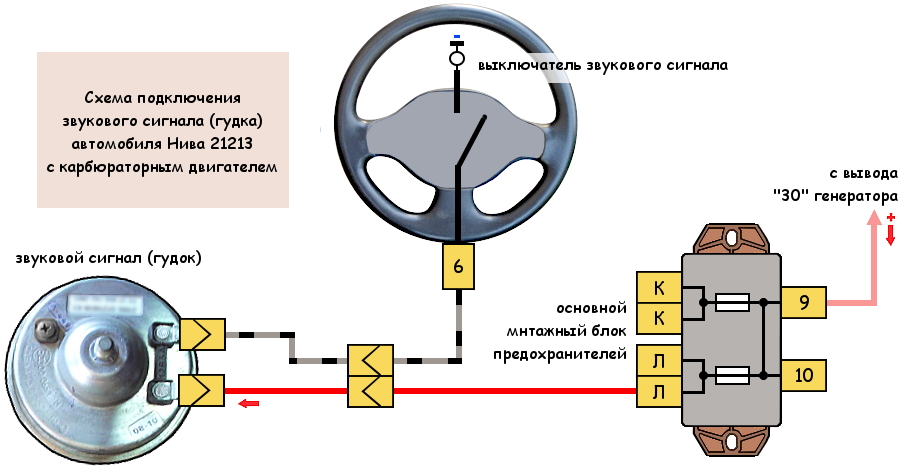 Поэтому было предположено, что белки, содержащие домен PH, действуют как факторы нуклеации, пространственно направляя активные сигнальные каскады во время хемотаксиса.
Поэтому было предположено, что белки, содержащие домен PH, действуют как факторы нуклеации, пространственно направляя активные сигнальные каскады во время хемотаксиса.
Неожиданно мы обнаружили, что ACA сильно обогащена в задней части хемотаксирующих клеток, а также на динамических внутриклеточных везикулах () (Kriebel et al. , 2003). Далее мы показали, что такое клеточное распределение необходимо для того, чтобы клетки выравнивались по типу «голова к хвосту» и текли во время хемотаксиса. В этом контексте мы предположили, что пространственное ограничение ACA в задней части клеток обеспечивает компартмент, из которого высвобождается цАМФ и притягивает клетки специфически в задней части клеток перед ними. Механизм, который регулирует секрецию цАМФ, еще предстоит определить, хотя установлено, что сама по себе АСА не является переносчиком цАМФ (Pitt 9).1031 и др. , 1992). Мы полагаем, как и другие до нас, что везикулярный экзоцитоз может регулировать секрецию цАМФ (Maeda & Gerisch, 1977).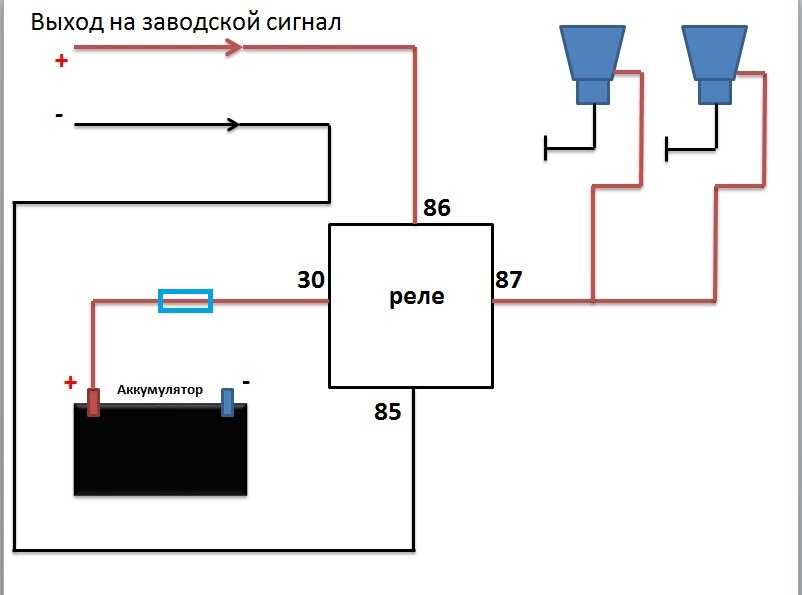 Мы показали, что хотя PI3K и CRAC необходимы для активации ACA, они не нужны для его клеточного распределения (Kriebel et al. , 2003; Comer & Parent, 2006). Таким образом, эти данные свидетельствуют о том, что различные механизмы независимо регулируют активность и клеточное распределение ACA. Поскольку существует значительный лаг между транслокацией CRAC и активацией ACA (см. выше), остается возможным, что активация ACA инициируется в передней части клеток и что дополнительные факторы, такие как TORC2, вовлекаются в процесс позже.
Мы показали, что хотя PI3K и CRAC необходимы для активации ACA, они не нужны для его клеточного распределения (Kriebel et al. , 2003; Comer & Parent, 2006). Таким образом, эти данные свидетельствуют о том, что различные механизмы независимо регулируют активность и клеточное распределение ACA. Поскольку существует значительный лаг между транслокацией CRAC и активацией ACA (см. выше), остается возможным, что активация ACA инициируется в передней части клеток и что дополнительные факторы, такие как TORC2, вовлекаются в процесс позже.
Исследования, проведенные на клетках хемотаксиса Dictyostelium , предоставили бесценную информацию о том, как клетки преобразуют, казалось бы, простые сигналы, такие как повышение уровня цАМФ, в сложные биологические реакции. Эти клетки делают это, тщательно ограничивая клеточное распределение ключевых компонентов сигнального каскада. Примечательно, что большая часть этой пространственной организации сохраняется, поскольку нейтрофилы человека также перераспределяют белки, содержащие домен PH, к своему переднему краю (Servant и др. , 1999; Слуга и др. , 2000; Ли и др. , 2003; Сюй и др. , 2003; Лакалль и др. , 2004; Нисио и др. , 2007). Мы предполагаем, что передача сигналов также будет пространственно ограничена у высших эукариот. Действительно, в нейтрофилах человека хемоаттрактанты также передают сигнал окружающим клеткам, стимулируя выработку и высвобождение дополнительных аттрактантов, таких как LTB 4 и интерлейкин 8 (IL-8), которые действуют аутокринно и паракринно, распространяя лейкоциты. хемотаксический ответ на окружающие клетки (Kuhns & Gallin, 1995; Кунс и др. , 2001; Йокомизо и др. , 2001). Тем не менее, было проделано очень мало работы, чтобы понять, как сложные пути ретрансляции сигналов регулируют миграцию и возвращение нейтрофилов. Недавно было показано, что дегрануляция нейтрофилов, опосредованная LTB 4 , защищает от цитомегаловирусной инфекции, но механизмы передачи сигнала, которые регулируют это, еще предстоит определить (Gaudreault & Gosselin, 2007).
, 1999; Слуга и др. , 2000; Ли и др. , 2003; Сюй и др. , 2003; Лакалль и др. , 2004; Нисио и др. , 2007). Мы предполагаем, что передача сигналов также будет пространственно ограничена у высших эукариот. Действительно, в нейтрофилах человека хемоаттрактанты также передают сигнал окружающим клеткам, стимулируя выработку и высвобождение дополнительных аттрактантов, таких как LTB 4 и интерлейкин 8 (IL-8), которые действуют аутокринно и паракринно, распространяя лейкоциты. хемотаксический ответ на окружающие клетки (Kuhns & Gallin, 1995; Кунс и др. , 2001; Йокомизо и др. , 2001). Тем не менее, было проделано очень мало работы, чтобы понять, как сложные пути ретрансляции сигналов регулируют миграцию и возвращение нейтрофилов. Недавно было показано, что дегрануляция нейтрофилов, опосредованная LTB 4 , защищает от цитомегаловирусной инфекции, но механизмы передачи сигнала, которые регулируют это, еще предстоит определить (Gaudreault & Gosselin, 2007).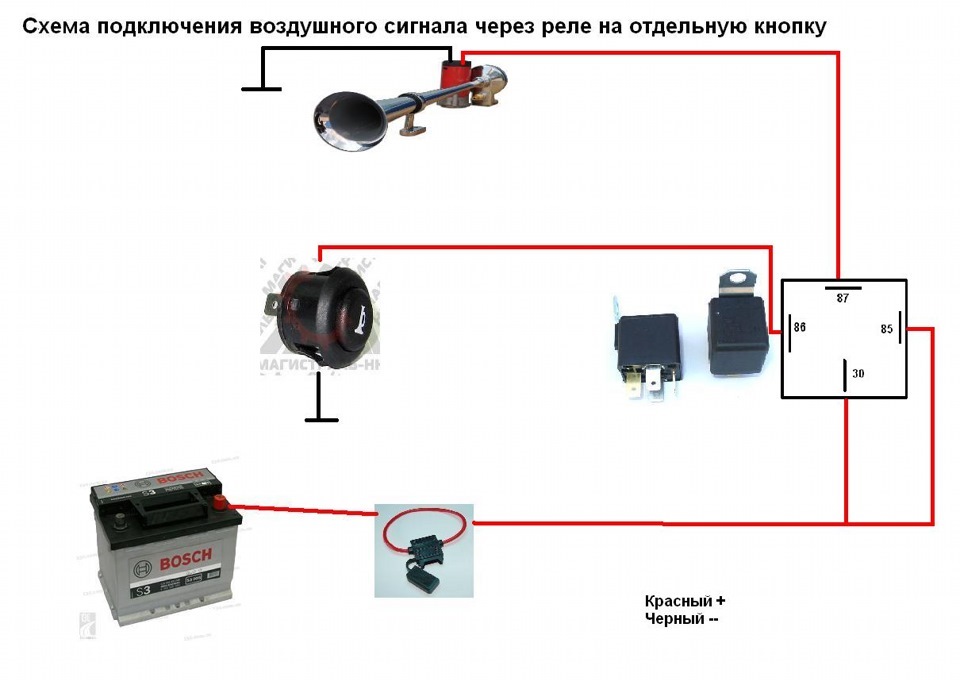 Будущая работа, несомненно, обнаружит новые и неожиданные открытия в комплексной регуляции ретрансляции сигналов во время миграции и развития клеток.
Будущая работа, несомненно, обнаружит новые и неожиданные открытия в комплексной регуляции ретрансляции сигналов во время миграции и развития клеток.
Мы благодарим г-на Пола Крибеля и д-ра Анну Баргоду за предоставление некоторых изображений, представленных в . Эта работа была поддержана Программой внутренних исследований Национальных институтов здравоохранения, Национального института рака, Центра исследований рака.
- Бадер С., Кортхольт А., Ван Хаастерт П.Дж. Семь фосфодиэстераз Dictyostelium discoideum расщепляют три пула цАМФ и цГМФ. Биохим Дж. 2007; 402:153–161. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Baggiolini M. Хемокины в патологии и медицине. J Интерн Мед. 2001; 250:91–104. [PubMed][Google Scholar]
- Багорда А., Михайлов В.А., Родитель К.А. Хемотаксис: двигаться вперед и держаться за прошлое. Тромб Хемост. 2006; 95:12–21. [PubMed] [Google Scholar]
- Baldauf SL. Глубокие корни эукариот. Наука. 2003; 300:1703–1706.
 [PubMed] [Google Scholar]
[PubMed] [Google Scholar] - Chen MY, Long Y, Devreotes PN. Новый цитозольный регулятор Пианиссимо необходим для опосредованной хемоаттрактантным рецептором и G-белком активации 12 трансмембранного домена аденилатциклазы в 9 клетках.1031 Диктиостелиум . Гены Дев. 1997; 11:3218–3231. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chisholm RL, Firtel RA. Взгляд на морфогенез из простой системы развития. Nat Rev Mol Cell Biol. 2004; 5: 531–541. [PubMed] [Google Scholar]
- Chisholm RL, Gaudet P, Just EM, Pilcher KE, Fey P, Merchant SN, Kibbe WA. dictyBase, база данных модельных организмов для Dictyostelium discoideum . Нуклеиновые Кислоты Res. 2006; 34:D423–427. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Comer FI, Lippincott CK, Masbad JJ, Parent CA. Опосредованная PI3K активация CRAC независимо регулирует активацию аденилатциклазы и хемотаксис. Карр Биол. 2005; 15: 134–139. [PubMed] [Google Scholar]
- Comer FI, Parent CA.
 PI 3-киназы и PTEN: как противоположности хемопритягиваются. Клетка. 2002; 109: 541–544. [PubMed] [Google Scholar]
PI 3-киназы и PTEN: как противоположности хемопритягиваются. Клетка. 2002; 109: 541–544. [PubMed] [Google Scholar] - Comer FI, Parent CA. Активность фосфоинозитид-3-киназы контролирует опосредованную хемоаттрактантом активацию и адаптацию аденилатциклазы. Мол Биол Селл. 2006; 17: 357–366. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Dinauer MC, MacKay SA, Devreotes PN. Циклический ретранслятор 3′,5′-AMP у Dictyostelium discoideum III. Взаимосвязь синтеза и секреции цАМФ во время сигнального ответа цАМФ. Джей Селл Биол. 1980а; 86: 537–544. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Dinauer MC, Steck TL, Devreotes PN. Реле циклического 3′,5′-АМФ у Dictyostelium discoideum V. Адаптация реакции передачи сигналов цАМФ во время стимуляции цАМФ. Джей Селл Биол. 1980b; 86: 554–561. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Eichinger L, Pachebat JA, Glockner G, et al. Геном социальной амебы Dictyostelium discoideum .
 Природа. 2005; 435:43–57. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Природа. 2005; 435:43–57. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Fey P, Gaudet P, Pilcher KE, Franke J, Chisholm RL. Dictybase и Dicty Stock Center. Методы Мол Биол. 2006; 346: 51–74. [PubMed] [Google Scholar]
- Фунамото С., Мейли Р., Ли С., Пэрри Л., Фиртел Р.А. Пространственная и временная регуляция 3-фосфоинозитидов с помощью PI 3-киназы и PTEN опосредует хемотаксис. Клетка. 2002;109: 611–623. [PubMed] [Google Scholar]
- Gaudreault E, Gosselin J. Опосредованное лейкотриеном B4 высвобождение противомикробных пептидов против цитомегаловируса зависит от BLT1. Вирус Иммунол. 2007; 20: 407–420. [PubMed] [Google Scholar]
- Herald D, Devreotes PN. Семейство рецепторов цАМФ Dictyostelium . Int Rev Cytol. 1992; 137Б: 35–47. [PubMed] [Google Scholar]
- Iijima M, Devreotes P. Опухолевой супрессор PTEN опосредует определение градиентов хемоаттрактанта. Клетка. 2002;109: 599–610. [PubMed] [Google Scholar]
- Инсалл Р.
 , Куспа А., Лилли П.Дж., Шаульский Г., Левин Л.Р., Лумис В.Ф., Девреотс П.Н. CRAC, цитозольный белок, содержащий домен гомологии с плекстрином, необходим для опосредованной рецептором и G-белком активации аденилатциклазы в Dictyostelium . Джей Селл Биол. 1994; 126:1537–1545. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Куспа А., Лилли П.Дж., Шаульский Г., Левин Л.Р., Лумис В.Ф., Девреотс П.Н. CRAC, цитозольный белок, содержащий домен гомологии с плекстрином, необходим для опосредованной рецептором и G-белком активации аденилатциклазы в Dictyostelium . Джей Селл Биол. 1994; 126:1537–1545. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Jin T, Zhang N, Long Y, Parent CA, Devreotes PN. Локализация комплекса G-белка b в живых клетках при хемотаксисе. Наука. 2000; 287:1034–1036. [PubMed] [Академия Google]
- Карник С.С., Гогонеа С., Патил С., Саад Ю., Такезако Т. Активация рецепторов, связанных с G-белком: общий молекулярный механизм. Тенденции Эндокринол Метаб. 2003; 14: 431–437. [PubMed] [Google Scholar]
- Кессин Р.Х. Диктиостелиум: эволюция, клеточная биология и развитие многоклеточности. Издательство Кембриджского университета; Cambridge: 2001. [Google Scholar]
- Kim JY, Borleis JA, Devreotes PN. Переключение программ развития и морфогенеза рецепторов хемоаттрактантов у Dictyostelium : подтипы рецепторов активируют общие реакции при различных концентрациях агонистов.
 Дев биол. 1998; 197: 117–128. [PubMed] [Google Scholar]
Дев биол. 1998; 197: 117–128. [PubMed] [Google Scholar] - Kimmel AR, Faix J. Генерация множественных нокаутных мутантов с использованием системы Cre-loxP. Методы Мол Биол. 2006; 346: 187–199. [PubMed] [Google Scholar]
- Kimmel AR, Firtel RA. Нарушение симметрии: регуляция развития Dictyostelium посредством хемоаттрактанта и ответной реакции морфогена. Curr Opin Genet Dev. 2004; 14: 540–549.. [PubMed] [Google Scholar]
- Kriebel PW, Barr VA, Parent CA. Локализация аденилатциклазы регулирует поток во время хемотаксиса. Клетка. 2003; 112: 549–560. [PubMed] [Google Scholar]
- Kriebel PW, Parent CA. Экспрессия и регуляция аденилатциклазы во время дифференцировки Dictyostelium discoideum . Жизнь ИУБМБ. 2004; 56: 541–546. [PubMed] [Google Scholar]
- Кульманн М., Попова Б., Неллен В. РНК-интерференция и антисмысловое замалчивание генов в Диктиостелиум . Методы Мол Биол. 2006; 346: 211–226. [PubMed] [Google Scholar]
- Kuhns DB, Gallin JI.
 Повышение уровня IL-8, ассоциированного с клетками, в экссудативных и обработанных A23187 нейтрофилах периферической крови человека. Дж Иммунол. 1995; 154:6556–6562. [PubMed] [Google Scholar]
Повышение уровня IL-8, ассоциированного с клетками, в экссудативных и обработанных A23187 нейтрофилах периферической крови человека. Дж Иммунол. 1995; 154:6556–6562. [PubMed] [Google Scholar] - Kuhns DB, Nelson EL, Alvord WG, Gallin JI. Фибриноген индуцирует синтез IL-8 в нейтрофилах человека, стимулированных формилметиониллейцилфенилаланином или лейкотриеном B(4) J Immunol. 2001; 167: 2869–2878. [PubMed] [Академия Google]
- Kuspa A. Мутагенез, опосредованный рестрикционными ферментами (REMI). Методы Мол Биол. 2006; 346: 201–209. [PubMed] [Google Scholar]
- Lacalle RA, Gomez-Mouton C, Barber DF, et al. PTEN регулирует подвижность, но не направленность во время хемотаксиса лейкоцитов. Дж. Клеточные науки. 2004; 117:6207–6215. [PubMed] [Google Scholar]
- Landree MA, Devreotes PN. Анализ хемотаксиса с использованием Dictyostelium discoideum в качестве модельной системы. Методы Мол Биол. 2004; 239:91–104. [PubMed] [Академия Google]
- Ли С.
 , Комер Ф.И., Сасаки А., Маклеод IX и др. Комплекс TOR 2 объединяет движение клеток во время хемотаксиса и передачу сигнала в Dictyostelium . Мол Биол Селл. 2005; 16: 4572–4583. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Комер Ф.И., Сасаки А., Маклеод IX и др. Комплекс TOR 2 объединяет движение клеток во время хемотаксиса и передачу сигнала в Dictyostelium . Мол Биол Селл. 2005; 16: 4572–4583. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Lee S, Parent CA, Insall R, Firtel RA. Новый Ras-взаимодействующий белок, необходимый для хемотаксиса и передачи сигнала циклического аденозинмонофосфата у Dictyostelium . Мол Биол Селл. 1999;10:2829–2845. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ли З., Ханниган М., Мо З. и др. Для направленного восприятия требуется G b-опосредованная PAK1 и зависимая от PIX альфа активация Cdc42. Клетка. 2003; 114: 215–227. [PubMed] [Google Scholar]
- Lilly PJ, Devreotes PN. Идентификация CRAC, цитозольного регулятора, необходимого для стимуляции гуаниновыми нуклеотидами аденилатциклазы у Dictyostelium . Дж. Биол. Хим. 1994; 269:14123–14129. [PubMed] [Google Scholar]
- Lilly PJ, Devreotes PN. Опосредованная хемоаттрактантом и ГТФгС стимуляция аденилатциклазы в Dictyostelium требует переноса CRAC на мембраны.
 Джей Селл Биол. 1995; 129:1659–1665. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Джей Селл Биол. 1995; 129:1659–1665. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Maeda Y, Gerisch G. Образование везикул в клетках Dictyostelium discoideum во время колебаний синтеза и высвобождения цАМФ. Разрешение ячейки опыта. 1977; 110: 119–126. [PubMed] [Google Scholar]
- Mahadeo DC, Parent CA. Сигнальное реле в течение жизненного цикла Dictyostelium . Curr Top Dev Biol. 2006; 73: 115–140. [PubMed] [Академия Google]
- Мейли Р., Эллсуорт С., Фиртел Р.А. Новая Akt/PKB-родственная киназа необходима для морфогенеза Dictyostelium . Карр Биол. 2000; 10: 708–717. [PubMed] [Google Scholar]
- Meili R, Ellsworth C, Lee S, Reddy TB, Firtel RA. Опосредованная хемоаттрактантом транзиторная активация и мембранная локализация Akt/PKB необходимы для эффективного хемотаксиса до цАМФ у Dictyostelium . EMBO J. 1999; 18:2092–2105. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Миллиган Г., Костенис Э. Гетеротримерные G-белки: краткая история.
 Бр Дж. Фармакол. 2006; 147 (Приложение 1): S46–S55. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Бр Дж. Фармакол. 2006; 147 (Приложение 1): S46–S55. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Muller-Taubenberger A. Применение флуоресцентных белковых меток в качестве репортеров в исследованиях визуализации живых клеток. Методы Мол Биол. 2006; 346: 229–246. [PubMed] [Google Scholar]
- Нисио М., Ватанабэ К., Сасаки Дж. и др. Контроль полярности и подвижности клеток с помощью фосфатазы PtdIns(3,4,5)P3 SHIP1. Nat Cell Biol. 2007; 9:36–44. [PubMed] [Google Scholar]
- Родитель CA, Blacklock BJ, Froelich WM, Murphy DB, Devreotes PN. События передачи сигналов G-белка активируются на переднем крае хемотаксических клеток. Клетка. 1998;95:81–91. [PubMed] [Google Scholar]
- Parent CA, Devreotes PN. Молекулярная генетика передачи сигнала у Dictyostelium . Энн Рев Биохим. 1996; 65: 411–440. [PubMed] [Google Scholar]
- Pitt GS, Milona N, Borleis JA, Lin KC, Reed RR, Devreotes PN. Структурно различные и стадиеспецифические гены аденилатциклазы играют разные роли в развитии Dictyostelium .
 Клетка. 1992; 69: 305–315. [PubMed] [Google Scholar]
Клетка. 1992; 69: 305–315. [PubMed] [Google Scholar] - Прабху Ю., Эйхингер Л. Dictyostelium репертуар семи рецепторов трансмембранных доменов. Eur J Cell Biol. 2006; 85: 937–946. [PubMed] [Google Scholar]
- Saran S, Meima ME, Alvarez-Curto E, Weening KE, Rozen DE, Schaap P. Передача сигналов цАМФ в Dictyostelium Сложность синтеза, деградации и обнаружения цАМФ. J Muscle Res Cell Motil. 2002; 23: 793–802. [PubMed] [Google Scholar]
- Сарбасов Д.Д., Али С.М., Сабатини Д.М. Растущая роль пути mTOR. Curr Opin Cell Biol. 2005;17:596–603. [PubMed] [Google Scholar]
- Sasaki AT, Chun C, Takeda K, Firtel RA. Локализованная передача сигналов Ras на переднем крае регулирует PI3K, полярность клеток и направленное движение клеток. Джей Селл Биол. 2004; 167: 505–518. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sasaki AT, Firtel RA. Регуляция хемотаксиса с помощью организованной активации Ras, PI3K и TOR. Eur J Cell Biol. 2006; 85: 873–895.
 [PubMed] [Google Scholar]
[PubMed] [Google Scholar] - Servant G, Weiner OD, Herzmark P, Balla T, Sedat JW, Bourne HR. Поляризация передачи сигналов рецептора хемоаттрактанта во время хемотаксиса нейтрофилов. Наука. 2000; 287:1037–1040. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Слуга Г., Вайнер О.Д., Нептун Э.Р., Седат Д.В., Борн Х.Р. Динамика рецептора хемоаттрактанта в живых нейтрофилах при хемотаксисе. Мол Биол Селл. 1999; 10:1163–1178. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Theibert A, Devreotes PN. Опосредованная поверхностными рецепторами активация аденилатциклазы у Dictyostelium . Регуляция гуаниновыми нуклеотидами в клетках дикого типа и мутантах с дефицитом агрегации. Дж. Биол. Хим. 1986; 261:15121–15125. [PubMed] [Академия Google]
- Вейер CJ. Dictyostelium морфогенез. Curr Opin Genet Dev. 2004; 14: 392–398. [PubMed] [Google Scholar]
- Williams HP, Harwood AJ. Полярность клеток и развитие Dictyostelium . Curr Opin Microbiol.