

Содержание
Грейферный механизм
Двухкривошипный
грейферный механизм киносъемочного
аппарата «Конвас-автомат«.
Гре́йферный
механи́зм, гре́йфер (от нем. greifen —
хватать) — разновидность скачкового
механизма в лентопротяжных
механизмах киносъёмочной, кинопроекционной, кинокопировальной аппаратуры
и в сканерах
для кинопленки,
служащий для прерывистого
перемещения кинопленки на
шаг кадра в фильмовом
канале и
имеющий по крайней мере один периодически
входящий в зацепление с перфорацией киноплёнки
и выходящий из него зуб.[1] По
сравнению с другими типами скачковых
механизмов (кулачковый, мальтийский,
пальцевый) грейферный механизм
обеспечивает наиболее высокую точность
перемещения кинопленки и получил
наибольшее распространение в киносъемочной
аппаратуре.
[Править]Принцип действия грейфера
Принцип
работы грейферного механизма основан
на преобразовании вращательного движения
вала привода в возвратно-поступательное
движение зуба по сложной траектории,
обеспечивающее четыре основных фазы
цикла: вход зуба в перфорацию, перемещение
кинопленки на шаг кадра, выход зуба из
перфорации и возврат в начало цикла.
Работа грейфера синхронизируется с
работой обтюратора таким
образом, чтобы после открытии обтюратора
кинопленка оставалось неподвижной до
его полного закрытия. Обычно оба механизма
имеют общий привод.
[Править]Рабочий угол
Рабочим
углом грейфера называется угол поворота
ведущего вала механизма, за который
происходит перемещение кинопленки на
шаг кадра. Отношение рабочего угла к
углу полного поворота ведущего вала
грейфера (как правило, 360°)
соответствует КПД грейфера.
Чем меньше рабочий угол грейфера, тем
дольше кинопленка остается неподвижной
и тем дольше обтюратор может быть открыт.
[Править]Основные требования
высокую
точность перемещения киноплёнки
(приводит к неустойчивости изображения
на экране по вертикали)В
современной киносъемочной аппаратуре,
предназначенной для съемок звуковых
фильмов, одно из важнейших требований
к грейферу — малошумность, поскольку
грейферный механизм является основным
источником звука.
Грейферный механизм должен иметь
высокий коэффициент полезного действия
— отношение времени покоя к периоду
полного рабочего цикла механизма. Эта
характеристика определяется рабочим
углом грейфера.Во время экспозиции или проекции (при
открытом обтюраторе) кинопленка должна
оставаться неподвижной, что требует
ее очень быстрого торможения после
перемещения. Для обеспечения минимальных
усилий при транспортировке кинопленки
и ее быстрой остановки, в некоторых
конструкциях используется временный
прижим кинопленки в фильмовом канале
на период покоя. Однако, наиболее
эффективным устройством для обеспечения
неподвижности кинопленки при открытом
обтюраторе, является контргрейфер.
(Контргре́йфер —
дополнительный механизм киноаппаратуры,
представляющий собой один или несколько
зубьев, входящих в перфорацию киноплёнки
в момент вывода из перфорации зуба
грейфера, и фиксирующий плёнку в
неподвижном положении. )

Классификация
Большинство
грейферных механизмов основано на
принципе кривошипа. Различают
Различают
кривошипно-кулисные, кривошипно-шатунные
и кулачковые грейферные механизмы. В
простейших любительских киносъемочных
аппаратах встречаются оригинальные
упрощенные конструкции грейфера.
Существуют однозубые и многозубые
грейферы. От количества зубьев грейфера
зависит нагрузка на перфорацию кинопленки,
поэтому многозубые грейферы применяются,
в основном, в кинопроекционной аппаратуре,
которая должна обеспечивать низкий
износ фильмокопий. В киносъемочной
аппаратуре многозубые грейферы не
получили широкого распространения
вследствие технологической сложности
изготовления многозубого грейфера с
высокой точностью, необходимой для
обеспечения устойчивости изображения.
Также грейферы бывают односторонними
и двухсторонними. Двухсторонние грейферы
имеют зубья, входящие в оба ряда
перфорации. Прецизионные грейферные
механизмы обеспечивают наивысшую
точность перемещения кинопленки и
применяются в специальных киносъемочных
аппаратах для комбинированных съемок,
рассчитанных на многократные экспозиции.
Некоторые из таких грейферов сконструированы
по оригинальным схемам, в том числе с
использованием пульсирующего фильмового
канала.
3. см. б.4, №3
БИЛЕТ 6
1.
После съемки кинофильма, его монтажа
и озвучения съемочная
группа получает негативы изображения
и фонограммы на отдельных пленках.
Фонограмма может быть как оптической
на кинопленке (в настоящее время при
изготовлении фильмов не применяется),
так и магнитной, нанесенной на кинопленку
или на отдельной магнитной ленте. Эти
пленки попадают на кинокопировальную
фабрику, где с них при помощи кинокопировальных
аппаратов производится
контактная или оптическая печать
дубльнегатива или непосредственно
всего тиража фильмокопий. После печати
все фильмокопии поступают в кинотеатры для
их демонстрации зрителям при
помощикинопроекторов.
2.
Киносъёмочный аппарат (кинока́мера,
КСА) — оптико-механическое устройство,
предназначенное для съёмки кинофильма.
При этом процесс съёмки связан с
получением на киноплёнке ряда
последовательных изображений фаз
движения объекта съёмки.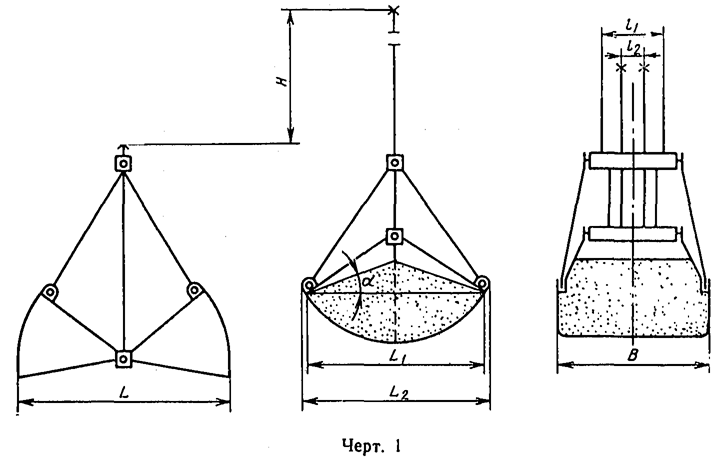
Первые
кинокамеры появились в конце XIX века с
изобретением кинематографа братьями
Люмьер.
До появления современных
электронных видеокамер кинокамеры
были единственным средством, позволявшим
производить запись движущегося
изображения для его последующего
воспроизведения кинопроектором на
экране. Как художественные,
так и документальные фильмы
снимались исключительно кинокамерами.
Для
записи движущегося изображения в
киносъёмочном аппарате применяется
прерывистое движение киноплёнки в
кадровом окне. В подавляющем большинстве
КСА для этого используется грейферный
механизм,
который представляет собой
кривошипно-кулисный или кривошипно-шатунный
механизм, совершающий возвратно-поступательное
движение грейфера — одного или
нескольких зубьев, входящих в контакт
с перфорацией киноплёнки
на время продвижения на расстояние,
равное шагу кадра, и выводимый из
перфорации во время холостого обратного
хода. Для предотвращения смазывания
изображения в момент продвижения плёнки
в КСА применяется обтюратор —
аналогфотографического
затвора,
перекрывающий световой поток от
съёмочного объектива к
плёнке. Главный тип обтюратора в
Главный тип обтюратора в
киносъёмочной технике — дисковый,
приводимый во вращение от вала грейферного
механизма. В аппаратах с лупой сквозной
наводки применяется зеркальный обтюратор,
расположенный под углом 45 градусов к
плоскости плёнки. В момент перекрытия
кадрового окна зеркальная поверхность
обтюратора направляет свет от объектива
на матовое
стекло коллективной
линзы аппарата, делая возможным
визирование непосредственно через
съёмочный объектив. В профессиональном
кинематографе кинокамеры для предотвращения
тряски изображения на экране устанавливают
на штатив,
который в свою очередь может крепиться
коператорской
тележке (dolly)
или устанавливаться на операторский
кран.
Специалист,
непосредственно работающий с кинокамерой
(управляющий ею) в процессе съёмки
кинофильма — кинооператор.
С появлением цифровых
кинокамер,
то есть видеокамер высокого разрешения,
предназначенных специально для
создания цифрового
кинофильма,
кинокамеры постепенно уходят в прошлое.
Основная
статья: Лентопротяжный
механизм (киноаппаратура)
Основная
статья: Грейферный
механизм
Основная
статья: Обтюратор
(оптика)
Основная
статья: Видоискатель#Сопряженный
визир киносъёмочного аппарата с
зеркальным обтюратором
В
киносъёмочной аппаратуре в качестве
привода используются электродвигатели
различного типа, пружинные механизмы
и ручной привод.
Счётчики
метража плёнки, переключатели и индикаторы
частоты киносъёмки (тахометры),
синхроотметчики, устройства записи на
кинопленку служебной информации
(например, временного
кода),
механизмы автоблокировки,
встроенные экспонометрические устройства,
звукопоглощающие боксы
3. Стереоскопическая
киносъемка
Киносъёмка, при
которой объект снимают одновременно с
двух или нескольких точек зрения так,
чтобы на киноплёнке (киноплёнках)
получались изображения, образующие
стереопары (См. Стереопара)
Стереопара)
(см. Стереоскопическое
кино).
Принципиальная схема С. к. однообъективным
киносъёмочным аппаратом (См. Киносъёмочный
аппарат)
с применением двух зеркал, установленных
под некоторым углом друг к другу, показана
на рис. 1, а. Объект рассматривается как
бы с двух точек зрения, расстояние между
которыми образует съёмочный базис
(Стереобазис).
В киноаппарате на киноплёнке фиксируются
два (левое и правое) изображения стереопары
(снимается стерсограмма). С. к. с
использованием двух объективов может
осуществляться либо двумя работающими
синхронно однообъективными киноаппаратами
на две раздельные киноплёнки, либо
двухобъективным стереоскопическим
киносъёмочным аппаратом (См. Стереоскопический
киносъёмочный аппарат)
на одну общую киноплёнку. Для изменения
величины стереобазиса двухобъективного
киноаппарата применяют систему раздвижных
зеркал (рис. 1, б). При
увеличении стереобазиса получают
стереоскопическое изображение с большей
пластикой, т.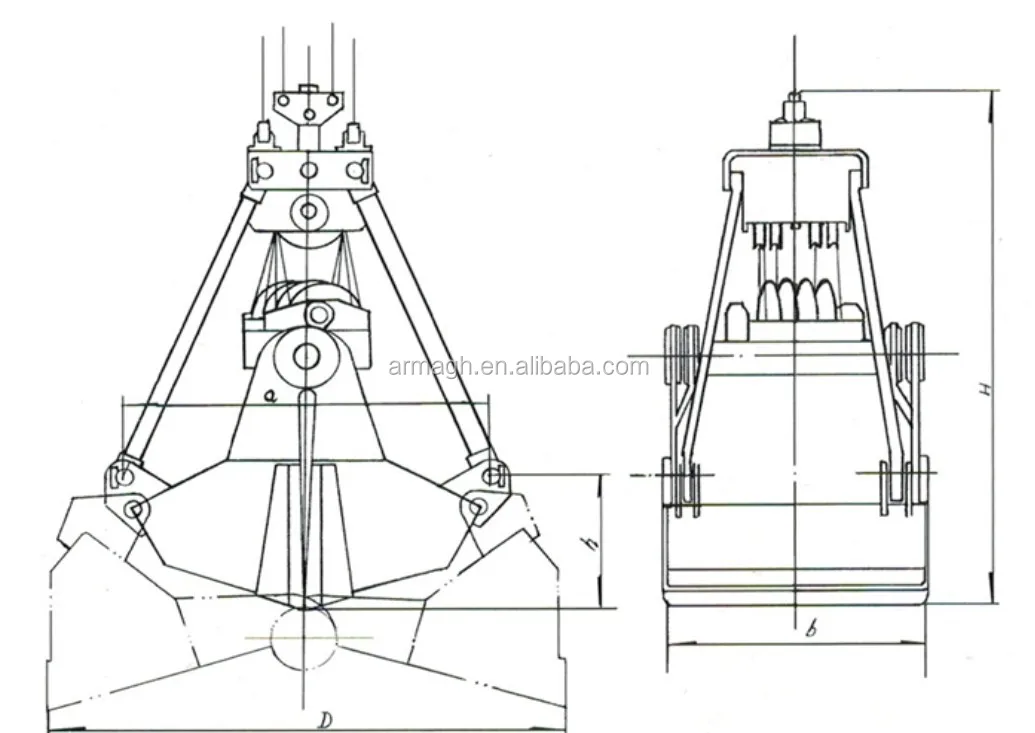 е. с более тонким различением
е. с более тонким различением
рельефа. Однако при проекции такое
изображение воспринимается зрителем
как макет, отличающийся от реального
объекта уменьшенными размерами.
Многостереопарная
киносъёмка обычно производится на общую
киноплёнку киноаппаратом с большим
числом (до 10) объективов (рис. 2). При
панорамограммной съёмке регистрируется
непрерывное изменение линейных Параллаксов
каждой точки изображения; при этом на
киноплёнке образуется как бы множество
переходящих одно в другое изображений
объекта, различающихся точками съёмки
(рис. 3). Такое изображение называется
«ксографией». Дальнейшее увеличение
пространственной информации об объекте
возможное применением его голограммы
(см. Голография).
БИЛЕТ 7
1.
2.
Обтюра́тор (фр. obturateur,
от лат. obturo —
закрываю) — механическое устройство
для периодического перекрывания
светового потока. Представляет собой
вращающийся секционированный диск, конус, цилиндр либо
двигающуюся возвратно-поступательно
шторку. Обтюраторы используются
Обтюраторы используются
в кинопроекторах, киносъемочных, кинокопировальных
аппаратах,
модуляционных радиометрах инфракрасного диапазона
и других оптико-механических
и фотоэлектрических приборах. В
инфракрасных радиометрах обтюратор
также может выполнять дополнительные
функции, например, в закрытом состоянии
— быть источником опорного излучения.
В
киносъемочном аппарате обтюратор
выполняет ту же роль, что и затвор в
фотоаппарате. Подвижная часть обтюратора
содержит прозрачные и непрозрачные
участки, которые попеременно располагаются
на пути светового потока. Работа
обтюратора синхронизируется со скачковым
механизмом таким
образом, чтобы кинопленка оставалась
неподвижной от начала открытия обтюратора
до его полного закрытия. Рассинхронизация
механизма, при которой кинопленка
движется при частично или полностью
открытом обтюраторе, называется «тягой
обтюратора».
Грейферные механизмы
Грейферные механизмы
Грейферные механизмы представляют собой особую разновидность рычажных и рычажно-оолиспастных грузозахватных устройств автоматического действия с различным исполнением привода рычажной системы.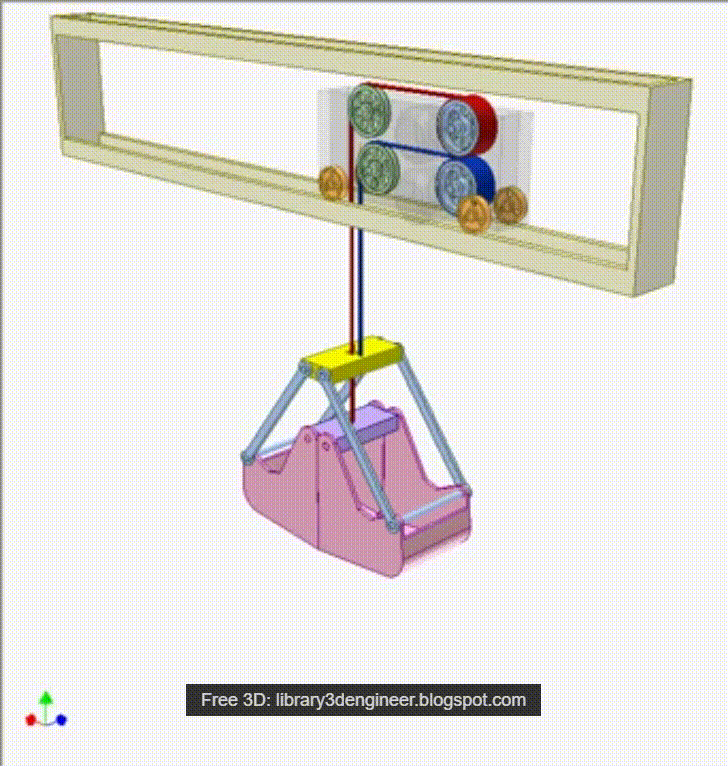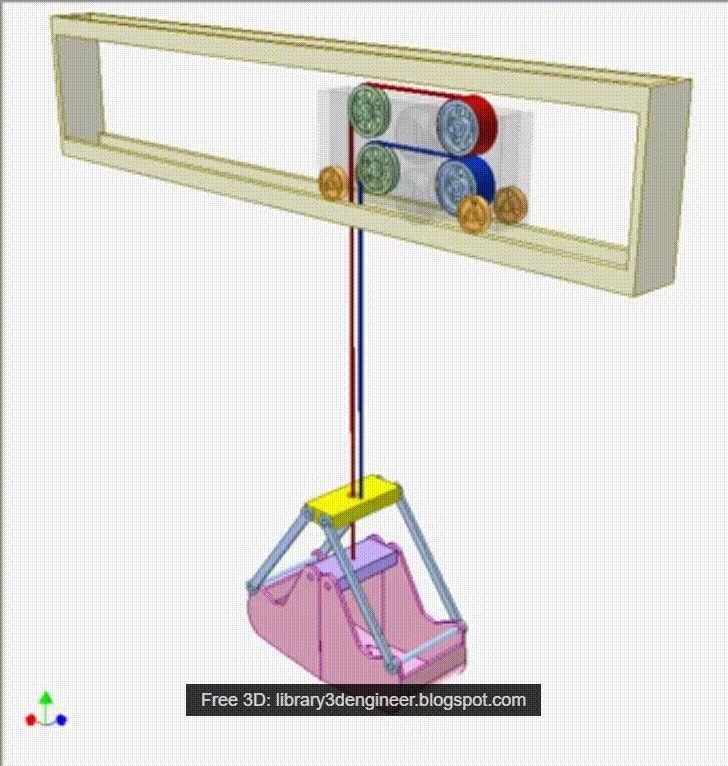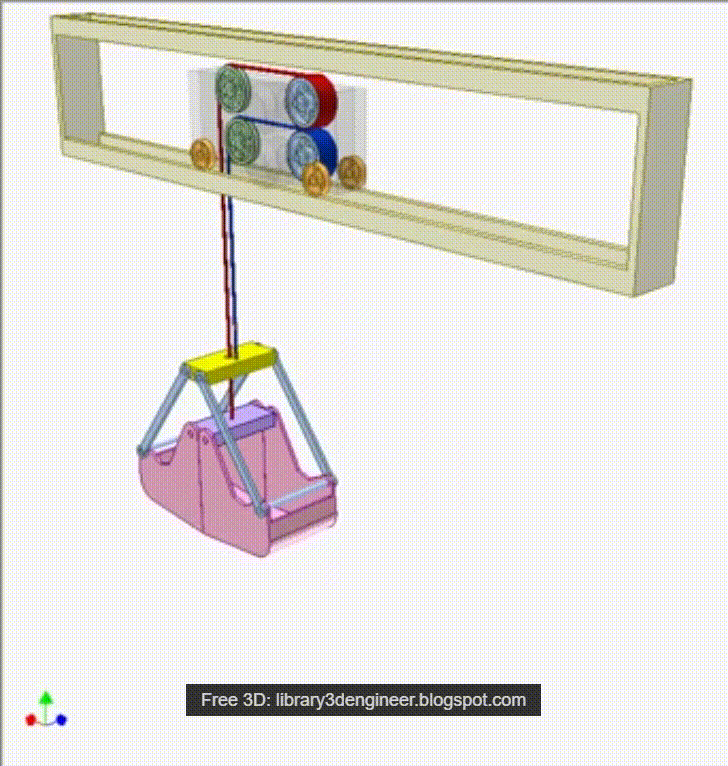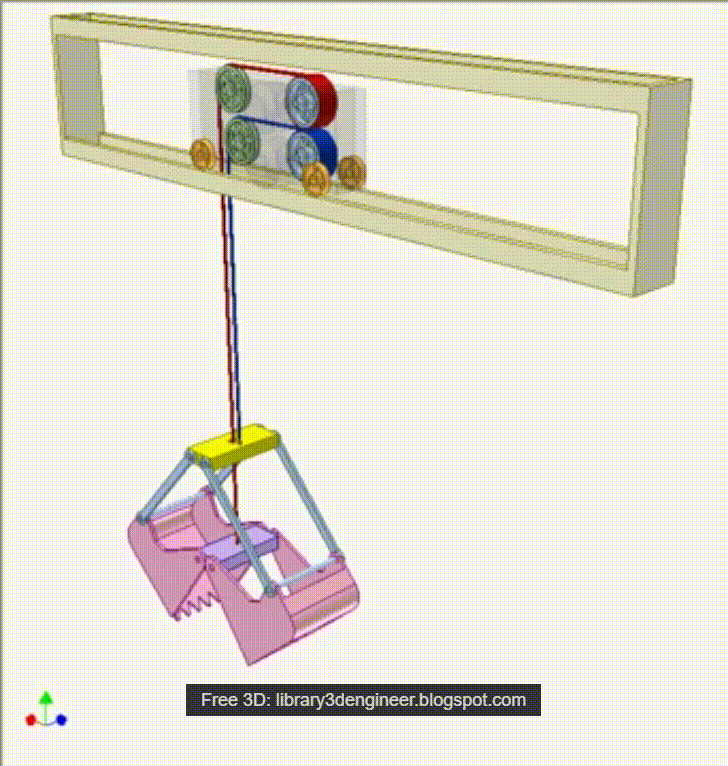 Основное назначение грейферных механизмов — перегрузка грузов, однородных в своей массе, например навалочных и круглого леса.
Основное назначение грейферных механизмов — перегрузка грузов, однородных в своей массе, например навалочных и круглого леса.
Известно несколько видов классификации грейферных механизмов: по конструктивным признакам, по технологическим признакам, по способу зачерпывания, по свойствам перегружаемых грузов.
По конструктивному признаку, в основе которого положен привод механизма раскрытия челюстей, грейферы подразделяются на канатные и приводные.
Рекламные предложения на основе ваших интересов:
Дополнительные материалы по теме:
Канатные грейферы, имеющие гибкую подвеску, приводятся в действие от механизма подъема крана, конструкция которого определяет тип грейфера. Известны грейферы: одно-, двух-, трех- и четырехканатные.
Однопалатные грейферы используются на кранах с одноба-рабанной лебедкой, не предназначенных для работы в грейферном режиме, поэтому несущий канат выполняет функцию замыкающего путем соединения с канатом, запакованным в грейфере. В результате зачерпывание осуществляется с помощью несущего каната, а раскрытие — с помощью специальных устройств на грейфере. В зависимости от конструктивного исполнения эти устройства позволяют раскрывать грейфер на любой высоте, на определенной высоте, при упоре о груз.
В результате зачерпывание осуществляется с помощью несущего каната, а раскрытие — с помощью специальных устройств на грейфере. В зависимости от конструктивного исполнения эти устройства позволяют раскрывать грейфер на любой высоте, на определенной высоте, при упоре о груз.
Двухканатные грейферы используются на кранах, предназначенных для работы в грейферном режиме. Они могут иметь: при двух канатах — один поддерживающий, другой замыкающий; при трех канатах— один поддерживающий и два замыкающих (встречаются очень редко), при четырех канатах — два поддерживающих и два замыкающих. Больше четырех канатов имеется только у специальных кранов большой грузоподъемности.
Увеличение числа канатов связано со стремлением уменьшить диаметр каната, а следовательно, диаметр блоков и направляющих.
У двухканатных грейферов канат образует одинарный полиспат, а у четырехканатных — сдвоенный.
По числу челюстей грейферы подразделяются на двух- и многочелюстные. Двухчелюстные могут быть жесткими, в этом случае все тяги имеют общую верхнюю ось, и рычажными, у которых верхние концы тяг шарнирно связаны с верхней траверсой. Жесткие типы грейферов не дают большого усилия для зачерпывания, и поэтому их применяют редко при перегрузке лег-козачерпываемых грузов.
Жесткие типы грейферов не дают большого усилия для зачерпывания, и поэтому их применяют редко при перегрузке лег-козачерпываемых грузов.
Рис. 1. Схема двухканатного грейфера:
1 — подъемный канат; 2— замыкающий канат.
Положение грейфера:
а — опускание на груз; б — зачерпывание; в — подъем; г — раскрытие
Многочелюстные подразделяются на грейферы с зависимой системой подвески, при которой замыкающий канат последовательно огибает блоки всех челюстей, и с независимой, при которой каждая челюсть имеет свой привод.
Раскрытие и закрытие челюстей у двухканатных (четырех-канатных) грейферов производятся на любой высоте подъема за счет специальных лебедок механизма подъема крана.
Приводные грейферы (моторные) подвешиваются к крану на гибкой связи, обеспечивающей выполнение операций по подъему, опусканию и переносу в пространстве. Челюсти закрываются и раскрываются специальными агрегатами, которые могут быть электрическими или электрогидравлическими, устанавливаемыми либо на верхней траверсе грейфера, либо на специальной траверсе с подводом электропитания к приводу с помощью кабеля. Приводные грейферы предназначены для перегрузки тяжелых и крупнокусковых навалочных грузов, а также круглого леса. Особенностью такого типа грейферов являются значительные внедряющие усилия на кромках челюстей.
Приводные грейферы предназначены для перегрузки тяжелых и крупнокусковых навалочных грузов, а также круглого леса. Особенностью такого типа грейферов являются значительные внедряющие усилия на кромках челюстей.
По технологическому признаку, т. е. по способу использования, грейферы подразделяются на нормальные, применяемые на перегрузке основной массы грузов, и специальные, применяемые только на определенных операциях перегрузочного процесса (штивующие) или на перегрузке отдельных видов грузов (чугун, крупнокусковые, металлолом, лес), к которым можно отнести грейферы с приводом раскрытия челюстей от механизма подъема крана и специальным приводом (моторные).
По способу зачерпывания грейферы могут быть подразделены на выполняющие эти операции без интенсифицирующих устройств и с интенсифицирующими устройствами. К первой группе относятся грейферы с фиксированной траекторией челюстей, гравитационные и якорные, ко второй — вибрационные и напорные.
По свойствам перегружаемых грузов классифицируются нормальные двухчелюстные грейферы. Классификация грейферов по этому признаку находится в прямой зависимости от транспортно-перегрузочных свойств основных массовых навалочных грузов (крупнокусковые, мелкокусковые, зернообразные и порошкообразные).
Классификация грейферов по этому признаку находится в прямой зависимости от транспортно-перегрузочных свойств основных массовых навалочных грузов (крупнокусковые, мелкокусковые, зернообразные и порошкообразные).
Основной характеристикой грейфера является его объем, который подбирается из условия использования наибольшей зачерпывающей способности, равной грузоподъемности крана, за вычетом соответственно веса грейфера.
Исследования, проведенные в Ждановском металлургическом институте и в Черноморском ЦПКБ, свидетельствуют, что производительность грейферного крана зависит от целого ряда факторов.
На основе этих факторов Черноморское ЦПКБ разработало сетку типоразмеров для угольных и рудных грейферов, а также для перегрузки стройматериалов и химикатов навалом применительно к грузоподъемности кранов 10; 12,5; 15 и 16 тс. В таблицах указаны емкость грейфера, вес грейфера, груза и грейфера вместе с грузом, марка груза с указанием размера кусков и их процентным содержанием в общей массе.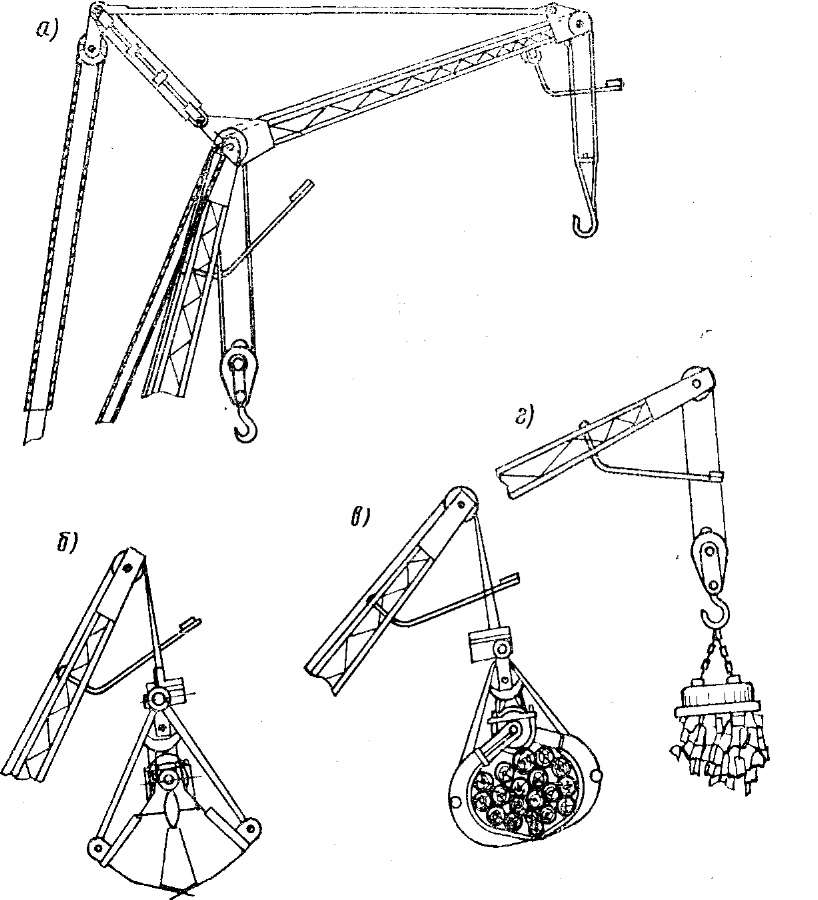 Кроме этого, имеются таблицы геометрических параметров грейферов. Однако в настоящей книге эта сетка грейферов не приводится ввиду ее большого объема.
Кроме этого, имеются таблицы геометрических параметров грейферов. Однако в настоящей книге эта сетка грейферов не приводится ввиду ее большого объема.
Рис. 2. Электромоторный виброгрейфер модели ВМГ-5:
а — порожний грейфер; б—д — перегрузочные операции, выполняемые грейфером.
Электроприводные грейферы предназначены для перегрузки труднозачерпываемых грузов, таких, как круглый лес, камень п др. Их изготовляют талевыми, винтовыми, секторными и лебедочными. В результате исследований существующих электроприводных грейферов для перегрузки леса, проведенных Московским лесотехническим институтом (МЛТИ) был выявлен ряд естественных недостатков. Установлено, что форма челюстей, размещение масс грейфера и система полиспаста не соответствуют характеру сопротивлений, преодолеваемых при зачерпывании. Изменение только конструкции не позволяет повлиять резко на характеристики, так как на зачерпывание леса из штабеля требуются значительные усилия при проникновении в толщу груза и на последующее подгребание.
Увеличение зачерпывающей способности, при изменении также формы челюстей, было достигнуто за счет применения эффекта от вибрации, разрушающей плотно лежащие ряды бревен в штабеле и способствующей интенсивному внедрению даже при малом весе грейфера. Вибратор устанавливается на челюстях или траверсах грейфера. На этом принципе построен электромоторный вибрационный грейфер модели ВМГ-5, разработанный МЛТИ. Грейфер (рис. 2) состоит из двух шарнирно соединенных челюстей серповидной формы со сплошным ножом. Форма челюстей выбрана таким образом, что в момент зачерпывания леса они занимают вертикальное положение. Под действием собственного веса и вибрации происходит заглубление челюстей в толщу леса. Смыкание челюстей после заглубления осуществляется полиспастом. Такой грейфер предназначен для выгрузки и погрузки круглого длинномерного леса в полувагоны, погрузки барж и штабелирования. Грузоподъемность грейфера 7 м3, при длине леса 6,5 м, площадь зева при сомкнутых челюстях 1,2 м2.
Рис. 3. Электрогидравлический грейфер для короткомерного баланса:
3. Электрогидравлический грейфер для короткомерного баланса:
а — выгрузка из трюма; 6 — погрузка платформ
Электрогидравлический грейфер для перегрузки короткомерного баланса, применяемый в порту Дюнкерк (Франция) представляет собой (рис. 46, а) две шарнирно связанные челюсти из полосовой стали с общей режущей кромкой. Каждая челюсть состоит из трех полос, соединенных ребрами жесткости с размещенными на них блоками. Смыкание и размыкание челюстей производятся гидроцилиндром, расположенным над шарнирной осью их соединения. Форма челюстей выбрана из условия эффективного зачерпывания леса из штабеля. Головка грейфера представляет собой агрегат, в котором расположена электро- и гидравлическая аппаратура, обеспечивающая работоспособность грейфера. Питание агрегата — от гибкого кабеля.
Отличительная особенность этого грейфера — наличие трех ветвей стального каната, запасованного через блоки челюстей, которые обеспечивают надежное обжатие балансов независимо от зачерпнутого объема при смыкании челюстей встык без создания переменной емкости. Грейфер применяется при перегрузке баланса длиной 1 — 1,22 м по прямому варианту — из судна на специальные железнодорожные платформы с высокими торцевыми бортами. Зачерпывает грейфер в трюме одновременно 5 мг. После укладки -на платформу специальными тележками производится торцовка балансов подбивкой к противоположному высокому борту. В местах выгрузки применяются напольные погрузчики, оборудованные аналогичными гидравлическими грейферами емкостью 2,5 ж3, с помощью которых они штабелируют баланс сплошными рядами.
Грейфер применяется при перегрузке баланса длиной 1 — 1,22 м по прямому варианту — из судна на специальные железнодорожные платформы с высокими торцевыми бортами. Зачерпывает грейфер в трюме одновременно 5 мг. После укладки -на платформу специальными тележками производится торцовка балансов подбивкой к противоположному высокому борту. В местах выгрузки применяются напольные погрузчики, оборудованные аналогичными гидравлическими грейферами емкостью 2,5 ж3, с помощью которых они штабелируют баланс сплошными рядами.
Рис. 4. Электрогидравлический грейферный захват СЕМ-160: а — в раскрытом состоянии; б — с грузом
Электрогидравлический грейферный захват модели СЕМ-160, изготавливаемый во Франции (рис. 4). Электрогидравлический привод, обеспечивающий раскрытие и закрытие клыков, расположен в верхней части захвата в металлическом кожухе. Кожух охлаждается вентилятором. Приборы расположены на поверхности диаметром 1300 мм, перекрытой разбирающимся на три части защитным капотом. Вес захвата 1100 кг. Максимальная грузоподъемность 10 тс. Время раскрытия клыков 1,5 сек, закрытия — 6 сек. Давление рабочей жидкости в гидросистеме 135 кГ/см2.
Вес захвата 1100 кг. Максимальная грузоподъемность 10 тс. Время раскрытия клыков 1,5 сек, закрытия — 6 сек. Давление рабочей жидкости в гидросистеме 135 кГ/см2.
Рис. 5. Двухчелюстной грейфер модели 2ГК-5-1
Двухчелюстной грейфер модели 2ГК-5-1, разработанный ЦПКБ МРФ, предназначен для перегрузки крупнокускового камня с использованием кранов грузоподъемностью 5 тс. Грейфер состоит из двух совковых челюстей, жестко связанных с рычажно-полиспастной системой. Конфигурация челюстей обеспечивает зачерпывание груза путем сгребающих движений (вместо внедрения под углом у обычных грейферов) при значительных усилиях, достигаемых за счет полиспастной системы кратностью, равной восьми. Поворот челюстей относительно верхнего (единственного) шарнира конструктивно выполнен таким образом, что при зачерпывании заклинивание камней между ними исключается. Замыкающий канат крана соединен с канатом, запасованным в полиспастной системе грейфера. Вытяжка каната составляет 10,5 м ввиду большой кратности полиспаста. Поддерживающий канат соединен с помощью скобы с двумя канатами, закрепленными снаружи к челюстям. Емкость грейфера 1,6 мг при весе 3300 кг.
Поддерживающий канат соединен с помощью скобы с двумя канатами, закрепленными снаружи к челюстям. Емкость грейфера 1,6 мг при весе 3300 кг.
Габариты грейфера: раскрытого 4300×1560×1820 мм, закрытого 2480X2280X1820 мм. Изготовитель грейфера — Балаковский судоремонтный завод МРФ. Производительность перегрузки камня грейфером в сравнении с работой ковшами (с заполнением вручную) увеличивается в 4,4 раза.
В Киевском речном порту грейфер был модернизирован с целью использования на перегрузке чугуна в чушках. Была уменьшена ширина, в результате чего емкость уменьшилась до 1,1 м3. Режущие кромки челюстей выполнены из высококачественной стали. Форма режущих кромок делается из сплошного листа, который затем разрезают по размеченной волнообразной кривой. Плотность прилегания челюстей, выполненная таким образом, исключает выпадание чушек при подъеме. Вес грейфера уменьшился на 400 кг, вес подъема увеличился на 200 кг.
Рис. 6. Многочелюстной грейфер для металлолома:
а — в раскрытом состоянии; б — с грузом металлолома
Производительность перегрузки модернизированным грейфером была достигнута 390 т в смену против 250 г в смену ковшами, число рабочих сократилось с 8 до 4.
Много челюстные канатные грейферы относятся к грейферам специального назначения, так как они обычно предназначаются для перегрузки определенных видов грузов. Они имеют значительный собственный вес, высокую кратность полиспаста — шести- или восьмикратные, сдвоенные и с зависимым движением челюстей. Число челюстей у существующих грейферов от 4 до 8.
Многочелюстной грейфер для металлолома, созданный конструкторским бюро Находкинского морского порта, состоит из восьми коробчатых челюстей, сваренных из листовой стали толщиной 36 мм. На концах челюстей кованые зубья. Челюсти соединены с тягами и нижней траверсой с помощью шаровых шарниров, обеспечивающих разворот их в вертикальной плоскости на угол 8—10°. Наличие шаровых шарниров способствует свободному внедрению челюстей в толщу металлолома, не вызывая деформации грейфера. Тяги выполнены в виде коробчатых балок с большим моментом инерции, что предотвращает деформацию геометрической формы грейфера при ударах о комингсы люка.
В отличие от известных многочелюстных грейферов у данного грейфера челюсти не стыкуются, и в результате этого свободные между ними пространства способствуют созданию хороших условий для зачерпывания и размещения внутри бесформенных кусков металлолома. Нижняя траверса пустотелая. Свободное пространство в ней рассчитано на установку внут,ри специальной малогабаритной электромагнитной шайбы диаметром 600 мм для увеличения интенсивности зачерпывания. Объем грейфера 1,8 м3 при собственном весе 4,8 т. Высота грейфера в закрытом состоянии 3200 м, диаметр 2800 мм.
Нижняя траверса пустотелая. Свободное пространство в ней рассчитано на установку внут,ри специальной малогабаритной электромагнитной шайбы диаметром 600 мм для увеличения интенсивности зачерпывания. Объем грейфера 1,8 м3 при собственном весе 4,8 т. Высота грейфера в закрытом состоянии 3200 м, диаметр 2800 мм.
Опыт применения данного -грейфера .показал Высокую эффективность и надежность в работе.
Многочелюстной грейфер для асфальта, применяемый в Ждановском морском порту, состоит из четырех леиестко-обраэных челюстей. Емкость грейфера 5,5 м3, собственный вес 8,2 г, вес зачерпываемого груза 6,6 т. Грейфер применяется для погрузки асфальта в полувагоны, а также при выгрузке барж. Последний слой в баржах выгружается модернизированным подгребающим грейфером емкостью 3,5 м3 модели 490, изготавливаемым Ждановским СРЗ по проекту Черноморского ЦПКБ. Модернизация произведена увеличением кратности полиспаста с 3 до 6 и установкой зубьев на режущей кромке.
Канатные грейферы для круглого леса предназначены для перегрузки круглого длинномерного леса, перевозимого в транспортных средствах (судах, полувагонах) и складируемого в штабелях россыпью плотными рядами. В эксплуатационной практике морских и речных портов в настоящее время применяются специальные двухчелюстные четырехка-натные грейферы с переменной емкостью челюстей. В отличие от обычных двухчелюстных лесные грейферы вместо сплошных с торцовыми стенками челюстей имеют три или четыре лапы или усеченные конусные челюсти, обеспечивающие переменную емкость, для плотного обжатия разного сечения зачерпываемых пачек леса.
В эксплуатационной практике морских и речных портов в настоящее время применяются специальные двухчелюстные четырехка-натные грейферы с переменной емкостью челюстей. В отличие от обычных двухчелюстных лесные грейферы вместо сплошных с торцовыми стенками челюстей имеют три или четыре лапы или усеченные конусные челюсти, обеспечивающие переменную емкость, для плотного обжатия разного сечения зачерпываемых пачек леса.
Рис. 7. Многочелюстной грейфер для перегрузки асфальта
Емкость лесного грейфера, определяемая наибольшей площадью сечения в сомкнутом состоянии лап (челюстей), принимается из расчета длины леса, укладочно-го веса его и грузоподъемности крана, на котором предполагается его использовать. Соблюдение этого условия является обязательным при проектировании лесных грейферов во избежание перегрузки крана.
Рис. 8. Двухчелюстной грейфер для круглого леса:
а — в раскрытом состоянии; б — выгрузка леса из лихтера
Двух канатный двухчелюстной грейфер- для кранов грузоподъемностью 7—10 тс, разработанный и внедренный в Ленинградском морском порту, состоит из двух траверс, шарнирно соединенных тягами. Челюсти выполнены усеченными и конусными. Для синхронной работы челюстей верхние концы имеют рычажное устройство. Грейфер хорошо внедряется в штабель леса. Наличие сплошных челюстей, ширина которых составляет 0,3 длины бревен, обеспечивает плотное обжатие зачерпнутого пучка леса даже при неровной Укладке (непараллельности отдельных бревен в штабеле). Челюсти имеют незначительный нахлест концов, что создает ему переменную емкость для более плотного обжатия пучка. Однако, как показала практика, этого нахлеста недостаточно, и поэтому бывают случаи заклинивания бревен на концах челюстей, а при зачерпывании небольшого объема пучка возможно и выскальзывание бревен. Недостатком грейфера является и его плохая устойчивость в вертикальном положении. Несмотря на указанные недостатки, грейфер успешно применяется на перегрузке лихтерного леса.
Челюсти выполнены усеченными и конусными. Для синхронной работы челюстей верхние концы имеют рычажное устройство. Грейфер хорошо внедряется в штабель леса. Наличие сплошных челюстей, ширина которых составляет 0,3 длины бревен, обеспечивает плотное обжатие зачерпнутого пучка леса даже при неровной Укладке (непараллельности отдельных бревен в штабеле). Челюсти имеют незначительный нахлест концов, что создает ему переменную емкость для более плотного обжатия пучка. Однако, как показала практика, этого нахлеста недостаточно, и поэтому бывают случаи заклинивания бревен на концах челюстей, а при зачерпывании небольшого объема пучка возможно и выскальзывание бревен. Недостатком грейфера является и его плохая устойчивость в вертикальном положении. Несмотря на указанные недостатки, грейфер успешно применяется на перегрузке лихтерного леса.
Двухканатный трехлапый грейфер к кранам грузоподъемностью 5 тс, разработанный ЦПКБ МРФ, состоит из двух траверсов и двух челюстей, соединенных шарнирно с верхней траверсой тягами. Челюсти выполнены
Челюсти выполнены
трехлапыми и позволяют изменять емкость в зависимости от зачерпнутого пучка леса. Для одновременного смыкания челюстей на верхних концах тяг имеется синхронизирующее рычажное устройство. Грейфер широко применяется на перегрузке круглого длинномерного леса из полувагонов и штабеля в .морских и речных портах. К недостаткам его следует отнести: относительно большой коэффициент конструкции (отношение собственного веса грейфера к его геометрической емкости) , составляющий 2790 кГ/м2, а также перекрестное попадание бревен в зазоры между лапами при зачерпывании леса, лежащего неровно в штабеле.
Рис. 9. Трехлапый грейфер для круглого леса
Для использования на перегрузке леса длиной 2 м ПКБ УПМО ММФ разработана модификация данного грейфера. Грейфер имеет меньший вес, синхронизатор на нижней траверсе и четырехканатную подвеску. Кроме этого, предусмотрена установка сбоку челюстей дополнительных лап, которые при перегрузке длинномерного леса не монтируются. Грейфер новой модификации имеет: вес 2094 кг без дополнительных лап, с лапами — 2244 кг, полезную грузоподъемность при лесе длиной 2 м — 1000 кГ при площади просвета 0,82 м2.
Грейфер новой модификации имеет: вес 2094 кг без дополнительных лап, с лапами — 2244 кг, полезную грузоподъемность при лесе длиной 2 м — 1000 кГ при площади просвета 0,82 м2.
По проекту ПКБ УПМО в ММФ изготавливается модификация трехлапого грейфера к кранам грузоподъемностью 10 тс. Конструкция его выполнена аналогично грейферу ЦПКБ МРФ, но для синхронной работы челюстей на верхних концах тяг и челюстей установлены зубчатые секторы. Недостатком его является малая ширина однолапой челюсти.
Используя опыт проектирования и эксплуатации, ПКБ 5’ПМО разработало специальные лесные грейферы к кранам грузоподъемностью 5; 10 и 15 тс. Грейферы двухчелюстные и в двухканатном исполнении. В отличие от эксплуатируемых лесных грейферов у новых челюсти сплошные и у одной из «их имеется углубление, что создает небольшое изменение емкости и тем самым обеспечивает плотность обжатия пучка леса. Кроме этого, такая конструкция челюстей исключает возможность попадания и заклинивания бревен в зазор между лапами, что случается при работе с трехлапыми грейферами. Однако производство этих грейферо-в пока не освоено.
Однако производство этих грейферо-в пока не освоено.
Рис. 10. Специальные лесные грейферы к кранам грузоподъемностью 5; 10; 15 тс
Рис. 11. Четырехлапый грейфер для плотового леса
Двухканатный четырехлапый грейфер к кранам грузоподъемностью 20 тс, разработанный Ленинградским морским портом, состоит из основных узлов аналогично специальным лесным грейферам. Для синхронной работы верхние концы челюстей имеют зубчатые секторы. Каждая челюсть выполнена в виде двух изогнутых лап, что создает возможность изменения емкости (просвета) на небольшую величину. Из известных грейферов он имеет самый выгодный коэффициент конструкции, составляющий 1150 кГ/м2.
Грейфер применяется только для перегрузки плотового леса из воды. Преимущества использования его особенно ощутимы в ледовых условиях. Обхватывая пучок в воде, грейфер обкалывает своим весом лед вокруг пучка и освобождает его для безопасного подъема и переноса.
Рис. 12. Четырехлапый грейфер с прижимом для короткомерного баланса
Небольшой радиус изгиба концов свободной укладки балансов россыпью при одновременном раскрытии челюстей и движении прижима вниз. Синхронное раскрытие челюстей обеспечивается симметричным расположением зубчатых секторов в месте их соединения на нижней траверсе и рычажным устройством на
Синхронное раскрытие челюстей обеспечивается симметричным расположением зубчатых секторов в месте их соединения на нижней траверсе и рычажным устройством на
Четырехканатный четырехлапый грейфер с прижимом к кранам грузоподъемностью 10 тс разработан Балтийским ЦПКБ для погрузки короткомерного баланса длиной 1 и 2 м на суда россыпью при пакетном хранении на окладе в увя-зочных проволочных комплектах.
Конструкция грейфера (рис. 12) выполнена аналогично специальным лесным грейферам и состоит из двух траверс и двулапых челюстей. Концы лап, подобно грейферу для плотового леса, имеют специальную конфигурацию и при смыкании челюстей заходят одна в другую, создавая переменную емкость. Профиль челюстей выбран из условия захвата Увязанного пакета балансов, лап вызван необходимостью верхней траверсе, соединяющим тяги. Прижим предназначен для укладки баланса в расстил в штабеле за счет собственного веса и специально выбранного профиля нижней его кромки. Прижим перемещается в процессе раскрытия челюстей по направляющим с роликами, установленными на нижней траверсе грейфера.
Грейфер берет пакет с причала, куда он доставляется либо тыловым краном из штабеля, либо другими транспортирующими средствами. После незначительного обжатия пакета на причале грейфером рабочий снимает увязочный проволочный комплект. Затем пакет плотно обжимается грейфером и переносится в трюм. После опускания в месте укладки производится раскрытие грейфера, и баланс укладывается в расстил благодаря направленному перемещению челюстей и воздействию прижима, опускающегося под собственным весом.
Испытания опытного образца грейфера были произведены на балансе длиной 1 м в Ленинградском морском порту. В процессе опытной перегрузки баланса в трюм судна была достигнута производительность 300 м3 в смену. Баланс укладывался в просвете люка сплошными рядами параллельно диаметральной плоскости (ДП) судна.
В связи с развитием в последние годы пакетных перевозок короткомерных балансов в полужестких стропах от дальнейшего внедрения и связанной с этим доводкой конструкции грейфера с целью устранения выявленных недостатков отказались.
Ковш грейферный: назначение, область применения, классификация и устройство механизма
Оборудование
Просмотров 1.1к. Обновлено
Ковш грейферный неотъемлемый элемент для проведения погрузочных и разгрузочных работ особой сложности или больших объёмов. В строительстве, металлургии, при проведении земельных и сельскохозяйственных работ порой не обойтись без автоматизированных разгрузочно-погрузочных работ.
С целью облегчения человеческого труда и повышения производительности в XX-XXI веках стал широко применяться грейфер.
Грейферные ковши различаются по своему устройству и предназначению. Данная статья раскроет темы: что такое грейферный ковш, в каких областях применяется данное приспособление, модификации грейферов.
Содержание
- Назначение и особенности грейферного ковша
- Ковши классифицируются по следующим категориям
- Устройство и разновидности грейферных ковшей
- Приводные (моторные)
- Канатный привод
- Принцип работы двух канатного грейфера
- 3-х и 4-х канатные устройства
- Разновидности грейферных ковшей
- Технические характеристики грейферного ковша (стандартный 0,45 м³)
- Грейферный ковш в работе
Назначение и особенности грейферного ковша
Семейство грейферных — навесных крановых грузозахватных устройств — довольно обширно. Приспособления варьируются от универсальных до узкопрофильных — клещеобразных, вильчатых, вилочных, крюков и прочих.
Приспособления варьируются от универсальных до узкопрофильных — клещеобразных, вильчатых, вилочных, крюков и прочих.
Ковш является одним из типов грейферов и представляет собой съемный металлический черпак большого размера, который крепится к грузоподъемной технике разной мощности, например, к экскаватору или подъемному крану.
Ковши с горизонтально расположенным гидроцилиндром предназначены для погрузки сыпучих материалов. Модели с гидроцилиндром, расположенным вертикально, помимо функции погрузки способны осуществлять копательные работы: рыть траншеи, канавы, ямы.
Грейферные механизмы по грузоподъемности делятся на три типа.
- Для сыпучих материалов легкого уровня тяжести — подъема зерна, сахарного песка или угольной пыли.
- Для сыпучих материалов среднего уровня тяжести — работы с песком, гравием разной фракции или цементом.
- Тяжёлые грейферные установки используют при работе с рудой, ферросплавами или бокситом.
На фотографии ниже представлено специальное оборудование в виде грейферного ковша для экскаватора.
Ковши классифицируются по следующим категориям
- По свойствам сыпучих материалов. В зависимости от свойств сыпучего материала грейферные ковши могут иметь характерные различия. Приспособления бывают открытого, закрытого и полузакрытого типа. Механизм закрытого типа при смыкании створок не позволяет материалу высыпаться из ковша.
- По траектории захвата материала. Копательные механизмы имеют траекторию, предназначенную для углубления в почву. Их применяют при разработке породы и выполнении земельных работ, таких как рытье котлована под фундамент. Если требуется осуществить погрузку материала, лежащего на поверхности, применяют ковши с траекторией движения челюстей, позволяющей сгребать необходимый материал.
- По ширине размаха клещей. В продаже имеются модели с фиксированной и изменяющейся шириной размаха челюстей. При проведении работ вглубь, таких, как рытье колодца, скважины и других подобных работ используют копательные устройства с фиксированной шириной размаха, челюсти которых врезаются в грунт. Ширина размаха челюстей должна совпадать с шириной копаемого углубления.
- Механизм независимой работы челюстей. При работе с неоднородными материалами применяются грейферы с механизмом независимого смыкания. Такая конструкция востребована в случаях, когда челюсть встречает на своем пути препятствия, мешающие совершать захват синхронно с другой челюстью. Интегрированный пружинный механизм решает возникшую проблему. Второй способ решения проблемы — наличие индивидуального привода на каждой клешне.
- Вращающийся грейфер. Установка может быть оснащена ротором. При наличии роторного устройства грейфер способен вращаться вокруг своей оси и захватывать грузы в любом положении.
Устройство и разновидности грейферных ковшей
Ковш грейферный различается по устройству на приводный и канатный.
Приводные (моторные)
Движение и управление движущимися челюстями ковша осуществляется при помощи отдельного привода. Моторные грейферы подразделяются по типу привода.
- Электромеханические. Считаются простыми и в то же время надежными захватами. Рабочее усилие от электродвигателя на захваты передается с помощью передаточного механизма.
- Гидравлические. Оборудованы гидроцилиндрами с системой шлангов. На фото ниже представлен типичный образец навесного оборудования с гидроприводом.
- Пневматические. По принципу работы подобны гидравлическому варианту, за исключением того, что в системе используется не жидкость, а воздух. Особое применение устройства нашли в цехах, оборудованных пневмосетью. К плюсам относят компактность и простоту конструкции.
- Электромагнитные. Под влиянием электротока создается магнитное поле. На корпусе ковша и подвижном магнитопроводе расположены катушки так называемого возбуждения. При подаче электричества образуется единое магнитное поле. Это приводит к захватыванию материала подвижными челюстями. Разгрузка производится после отключения электричества. Челюсти открываются под собственным весом. Применяются на предприятиях, связанных с переработкой металлолома.
Моторные ковши отличаются возможностью быстрого монтажа, компактностью и большой универсальностью. При этом они имеют меньшую вместительность в сравнении с канатными. Для стабильной работы и минимизирования перегрузок оснащаются предохранителями.
Особое применение моторные грейферы нашли на экскаваторной технике. Ковш грейферный для экскаватора дает возможность создавать необходимое давление при работе с грунтом. При этом он может независимо от своей массы работать с плотными породами.
К основным недостаткам моторных грейферов относят необходимость прогрева гидравлической системы при низких температурах.
Канатный привод
Данный вариант выполняется в одно-, двух-, трех- и четырех канатном вариантах.
Одноканатный. Манипуляции производятся одним канатом, который отвечает и за подъем механизма, и за работу челюстей. Используется на кранах с одной лебедкой.
Среди достоинств — быстрый монтаж и простота в эксплуатации.
Принцип работы устройства происходит в следующей последовательности.
- Процесс опускания. Полностью открытый грейфер опускается до упора на основание рабочей поверхности. При необходимости ковш можно «бросить», чтобы более глубоко внедриться в твердую породу.
- Захват. Срабатывает механизм защелкивания, и начинается процесс подъёма. При подъёме, челюсти постепенно смыкаются.
- Подъем. Трос продолжает поднимать ковш на нужный уровень.
- Разгрузка. Ковш опускается на поверхность, касание приводит к открытию затворного замка. Последующий подъем механизма приводит к открыванию ковша и выгрузке материала, после чего устройство готово к повторному циклу.
Двухкомнатный ковш оснащен двумя независимыми тросами. Один осуществляет подъем и опускание ковша, второй отвечает за работу челюстей.
Принцип работы двух канатного грейфера
- Опускание. При опускании ковша трос, отвечающий за работу челюстей, ослаблен.
- Захват материала. При полном опускании на породу трос, отвечающий за работу челюстей, начинает двигаться верх. Это приводит к захвату породы.
- Подъем. После закрытия челюстей начинается подъем ковша. При подъёме нагрузка распределяется равномерно на оба троса, что минимизирует их износ. Для синхронной работы двух независимых тросов применяются синхронизирующие устройства.
- Разгрузка. Нагруженный ковш может быть выгружен на любой высоте. Для этого трос, отвечающий за работу челюстей, ослабляется. После выгрузки ковш готов к повторному циклу.
3-х и 4-х канатные устройства
Принцип работы 3-х и 4-х канатных устройств не сильно отличается от работы двух канатного. У трех канатной модели один канат по-прежнему отвечает за подъем ковша, а два остальных синхронно управляют челюстями.
У четырех канатного два каната считаются поддерживающими, а два других — управляющими механизмом погрузки и выгрузки.
Канатные грейферы, в отличие от приводных, отличаются более высокой производительностью за счет большего веса и размера ковша.
В то же время имеют ряд недостатков — быстро изнашиваются и не подходят для разработки плотных пород грунта. Для работы с канатными грейферами требуется наличие тросов, которые имеют свойство раскачиваться и закручиваться.
Чтобы решить проблему раскачивания и возможного закручивания канатов на кран устанавливают успокоители тросов. Канатные грейферы устанавливают не только на подъёмные краны, но и на экскаваторы.
Ковш грейферный для экскаватора выпускается трех типов: легкий, средний и тяжелый. Ковши подбираются зависимо от плотности породы, с которой им предстоит работать.
Если неправильно подобрать тип ковша, то при разработке особо плотных грунтов он может оказаться малоэффективным, так как ему не хватит нагрузки, чтобы глубоко врезаться в грунт.
Разновидности грейферных ковшей
Грейферные ковши отличаются наличием металлических челюстей, при помощи которых производится захват материала. В зависимости от количества челюстей устройство бывает:
- одночелюстное;
- двухчелюстное;
- многочелюстное (максимум восемь челюстей).
Многочелюстные механизмы нашли свое применение в условиях работы с трудно зачерпываемыми материалами (руда крупных фракций). Челюсти в таких грейферах выполнены в форме серпа.
Ковш грейферный на сегодняшний день может быть выполнен с достаточно широким спектром необходимых свойств. В зависимости от целей и требуемой производительности можно подобрать оптимальный вариант.
Технические характеристики грейферного ковша (стандартный 0,45 м³)
| Высота | м. | 1,75 |
| Наименование | Единицы измерения | характеристики |
| Ширина захвата | мм. | 1650 |
| Ширина режущей кромки | 850/650 | |
| Высота подъема | 3510 | |
| Объём ковша | м³ | 0,45 |
| Грузоподъемность | тонн | 0,4 |
| Масса ковша | кг. | 280/260 |
| Длина | м. | 0,84 |
| Ширина | 1,15 |
Грейферный ковш в работе
Грейферный механизм для киноаппарата
ОПИСАНИЕ
ИЗОБРЕТЕНИЯ
К АВТОРСКОМУ СВИДЕТЕЛЬСТВУ и1540244
Союз Советских
Социалистических
Республик (61) Дополнительное к авт. свид-ву (22) 3 аявлено 05.11.75 (21) 2187059/10 с присоединением заявки № (23) Приоритет
Опубликовано 25.12.76. Бюллетень № 47
Дата опубликования описания 02.02.77 (51) M. Кл, G ОЗВ 1/22
Государственный комитет
Совета Министров СССР (53) УДК 778.531 (088.8) ло делам изобретений и открытии
72) Авторы
ДЛЯ КИНОАППАРАТА
Изобретение относится к кинотехнике, а именно к устройствам для точного прерывистого транспортирования киноленты, например, в прецизионных киносъемочных и кинокопировальных аппаратах, Известны грейферные механизмы для прерывистого транспортирования киноленты, осуществляемого подвижными зубьями грейфера (2), (3).
Эти механизмы наряду с подвижными зубьями грейфера имеют также подвижные или неподвижные зубья контргрейфера, предназначенные для повышения точности прерывистого транспортирования киноленты.
Принцип работы контргрейфера заключается в том, что после окончания прерывистого передвижения грейфером киноленты ее перфорации, расположенные на одном уровне по обеим сторонам ленты, фиксируются зубьями контргрейфера по высоте и ширине. В одних конструкциях грейферных механизмов зубья контргрейфера сами входят в перфорации киноленты, а в других кинолента надевается на неподвижные зубья контргрейфера подвижным фильмовым каналом.
В грейферном механизме с подвижными зубьями контгрейфера между последними и направляющими втулками этих зубьев должен быть гарантированный зазор. Поэтому положение зуба контргрейфера относительно кадрового окна может несколько измениться, что приводит к погрешности при фиксировании киноленты. Грейферный механизм с неподвижными зубьями контргрейфера обеспечивает более высокую точность прерывистого транспортирования киноленты на шаг кадра, равную 0,005 — 0,01 мм.
Наиболее близким к предлагаемому является грейферный механизм, содержащий кадровую рамку с неподвижными зубьями контргрейфера и ведущий вал с двумя кулачками, один из которых взаимодействует с подвижными зубьями грейфера, а другой — с качающимся фильмовым каналом. Фильмовый канал попеременно надевает киноленту на зубья грейфера, прерывисто транспортирующего ее на шаг кадра, и на зубья контргрейфера, фиксирующего киноленту относительно кадрового окна во время экспонирования (1).
Размеры фиксирующего сечения зуба контргрейфера выполняются равными стандартным размерам перфорации. Однако вследствие усадки киноленты величина перфорационного отверстия всегда несколько меньше сечения зуба контргрейфера. Поэтому кинолента надевается на зубья контргрейфера всегда с некоторым натягом, вызывающим деформацию кромок перфораций и повышенное трение этих кромок о поверхно30 сти зубьев. Большая и неравномерная по пе54О244
60 риметру сечения зуба сила трения приводит к неодинаковому для различных перфораций распределению внутренних напряжений и величин деформации их кромок, что сии>кает точность расположения перфораций киноленты относительно кадрового окна и повышает неустойчивость экспонированного или копировального изображения.
Недостатком известных грейферных механизмов является повышенное трение кромок перфорации киноленты о поверхности зубьев контргрейфера вследствие усадки ленты, что приводит к преждевременному и повышенному износу зубьев контргрейфера и снижает точность прерывистого транспортирования киноленты.
Цель изобретения — повышение точности транспортирования киноленты.
Указанная цель достигается тем, что механизм снабжен связанным с контргрейферными зубьями ультразвуковым преобразователем, соединенным через усилитель с генератором, причем в электрическую цепь между усилителем и преобразователем включены контакты, взаимодействующие с введенным дополнительным кулачком, установленным на приводном валу, На чертеже представлена схема предлагаемого грейферного механизма киноаппарата.
Грейферный механизм киноаппарата содер>кит ведущий вал 1, на котором закреплен кулачок 2, связанный с помощью рамки 3 и рычагов 4, 5 с кулачком 6, помещенным в грейферную рамку 7, снабженную зубьями 8 и направляющими 9. На этом же валу установлен кулачок 10, который связан с помощью рамки 11 и рычагов 12, 13, с кулачком
14, расположенным в кронштейне 15 фильмового канала 16, установленного с возможностью поворота вокруг оси 17. Кроме того, на ведущем валу помещен дополнительный кулачок 18, имеющий возможность взаимодействия к контактной группой 19.
Грейферный механизм содержит кадровую рамку 20, в прорезь которой входят зубья 21 контргрейфера 22, генератор 23 и усилитель
24 электрических ультразвуковых колебаний, а также пьезоэлектрический преобразователь
25 ультразвуковых колебаний в механические, связанный с зубьями 21 контргрейфера 22 и с контактной группой 19.
Механизм работает следующим образом.
При вращении ведущего вала 1 кулачок
2, взаимодействуя с рамкой 3 рычагов 4, 5, при помощи кулачка 6 перемещает рамку 7 с зубьями 8 грейфера в направлении транспортирования киноленты 26 и обратно по направляющим 9. В это время кулачок 10 взаимодействует с рамкой 11 рычагов 12, 13, при помощи кулачка 14 поворачивает фильмовый канал 16 с кинолентой 26 вокруг оси 17. При
35 этом кулачок 18 взаимодействует с контактной группой 19, замыкая и размыкая цепь выходного сигнала генератора 23.
После надевания киноленты 26, находящейся в фильмовом канале 16, на зубья 8 грейферной рамки 7 последняя транспортирует киноленту (по стрелке А) на шаг кадра. Затем фильмовый канал 16, поворачиваясь вокруг оси 17, снимает киноленту 26 с зубьев
8 рамки 7 и надевает ее на неподви>кные зубья 21 контргрейфера 22. В это время кулачок 18 замыкает контакты группы 19, и электрические колебания ультразвуковой частоты с выхода генератора 23 поступают на усилитель 24, а затем на пьезоэлектрический преобразователь 25. Таким образом, при надевании перфораций киноленты на зубья 21 контргрейфера 22 в зубьях возбуждаются механические колебания ультразвуковой частоты порядка 50 — 100 кГц.
После полного надевания перфораций киноленты 26 на зубья 21 контргрейфера 22 кинолента экспонируется и снимается фильмовым каналом 16 с зубьев 21 контргрейфера 22, после чего кулачок 18 размыкает контактную группу 19 и вибрация зубьев контргрейфера прекращается. Затем цикл повторяется снова.
Вибрация зубьев 21 контргрейфера 22 при надевании на них перфораций киноленты уменьшает силу трения между зубьями и кромками перфораций. Это повышает равномерность деформации кромок по контуру перфораций и увеличивает в 2 — 2,5 раза точность транспортирования киноленты на шаг кадра.
Формула изобретения
Грейферный механизм для киноаппарата, содержащий приводной вал с кулачками, кинематически связанными соответственно с рамкой, несущей зуб грейфера, и подвижным фильмовым каналом, и кадровую рамку с зубьями контргрейфера, отличающийся тем, что, с целью повышения точности транспортирования киноленты, он снабжен связанным с контргрейферными зубьями ультразвуковым преобразователем, соединенным через усилитель с генератором, причем в электрическую цепь между усилителем и преобразователем включены контакты, взаимодействующие с введенным дополнительным кулачком, установленным на приводном валу.
Источники информации, принятые во внимание при экспертизе:
1. Мелик-Степаняна А. М. и др. «Детали и механизмы киноаппаратуры», «Искусство», М., 1959 г., стр. 228 (прототип).
2. Авт. св. СССР № 168593, М. Кл. G ОЗВ
1/22, 1963.
3. Авт. св. № !63488, М. Кл, G ОЗВ 1/22, 1962.
540244
Составитель С. Коврина
Техред Е. Петрова
Корректор Е. Хмелева
Редактор Т. Иванова
Типография, пр. Сапунова, 2
Заказ 3091/9 Изд. Мв 381 Тираж 575 Подписное
ЦНИИПИ Государственного комитета Совета Министров СССР по делам изобретений и открытий
113035, Москва, 5К-35, Раушская наб., д. 4!5
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
О разработках и технических решениях.
…
Пожалуй, не существует такой сферы деятельности, в которой не возникало бы специфических проблем. Исключением не становятся и работы по перевалке сыпучих и кусковых материалов с помощью грейферов. Пыль, недостаток обслуживания механизмов, уязвимости и повреждения в ходе работ — факторов, способных повлечь вред здоровью, простои и материальный ущерб — Вам это знакомо? В этой статье мы расскажем, какие разработки, способные справиться с решением подобных проблем, использует итальянская компания Negrini.
Система пылеподавления
Существует проблема, с которой сталкиваются все порты, занимающиеся перевалкой груза, особенно угля. Во время перевалочных работ с участием грейферов от сыпучих грузов и кусковых пород поднимаются клубы пыли. Они снижают видимость, чем создают препятствия рабочему процессу и наносят ущерб здоровью и экологии. Решить данную проблему способна встроенная в грейферы Negrini система пылеподавления, применимая к любому типу работ, в ходе которых в воздух поднимается большое количество пыли. Система пылеподавления грейферов Negrini основана на распылении жидкости с помощью форсунок. Принцип ее работы в том, что вода под высоким давлением распыляется с помощью сопел (форсунок) ультратонкого распыления непосредственно в рабочей области. Пыль под воздействием водного облака становится тяжелее и оседает, что делает воздух чище и безопаснее. Технология была разработана компанией Negrini специально для России, как для лидирующего по добыче угля государства.
Принцип работы системы подавления пыли
Система автоматической смазки
Каждому грейферу требуется определенное обслуживание, а именно — шприцевание шарнирных частей. Зачастую сделать это непросто, так как грейферы могут достигать высоты 5 метров, ввиду чего приходится прибегать к использованию лестниц и спецтехники для подъема работника — а это небезопасно и времязатратно.
Грейферы Negrini оснащены централизованной смазкой, что делает смазку безопасной и увеличивает срок службы грейфера. Работнику достаточно подать смазку в отведенный для этого канал, она разойдется по шлангам и смажет все места, даже труднодоступные. Кроме того, это дает ряд преимуществ перед ручной смазкой:
- Исключение поднятия человека на высоту, что уменьшает риск получить тяжелую травму;
- Повышение производительности, и как следствие — увеличение прибыли;
- Исключение фактора, потенциально способного привести к простоям за счет сокращения количества неисправностей, вызванных некачественным смазыванием;
- Снижение затрат на проведение ремонта и закупки запасных деталей;
- Точное дозирование уменьшает расход смазочных материалов, а также предотвращает загрязнение окружающей среды;
- Повышение безопасности на рабочем месте ввиду того, что смазочные материалы не капают на пол/землю, а значит вероятность поскользнуться мала;
- Труднодоступные места смазки и все точки трения объединяются и надежно смазываются, что также повышает безопасность.
Система автоматической смазки
Дополнительные крышки и накладки
Частой проблемой при повышенной запыленности является забивание полиспастов пылью и грязью, что приводит к частым ремонтам. Дополнительные крышки позволяют уменьшить попадание в подвижные механизмы грейфера пыли и частиц груза, что также увеличивает срок службы грейфера и позволяет значительно сократить простои из-за ремонтов.
Грейферный ковш с крышкой
При перевалке мелкодисперсного груза зачастую происходит высыпание материала из-за неплотного закрытия челюстей грейфера. Чтобы не допустить даже малейшего высыпания материала между челюстей, компанией Negrini были разработаны специальные быстросъемные накладки, которые не допускают высыпания груза, закрывая щель между челюстями.
Дополнительные накладки
Сменные зубья Esco
Очень часто грейферы не просто используются для перевалки насыпного груза, но и копают вгрызаясь в различные породы. В таких случаях, зубья незаменимы. Однако в свою очередь они являются одной из наиболее подверженных износу частей. По этой причине компания Negrini использует в конструкции собственных грейферных ковшей широко доступные и высококачественные сменные зубья Esco, которые являются оптимальным сочетанием износостойкости и способности внедрения в сложных условиях. Зубья изготовлены из высокопрочной стали Hardox, кроме того — они сменные, что позволяет легко заменять пришедшие в непригодность на новые. Для работы с наиболее сложными грузами возможно использование длинных зубьев, называемых “рыхлителями”.
Грейфер с зубьями Esco
Защита гидроцилиндров от повреждений
В практике, к сожалению, часто встречается ситуация, когда операторы перегружателей и экскаваторов допускают ошибку, и по неосторожности в условиях ограниченного пространства происходит удар грейфера, например, о вагон. В случаях, если грейфер гидравлический или электрогидравлический, возможны повреждения гидроцилиндров, выход из строя которых недопустим и приводит к дорогостоящему и времязатратному ремонту. Во избежание таких серьезных поломок были предусмотрены элементы защиты гидроцилиндров от повреждений — защитные стальные кожухи гидроцилиндров, которые принимают удары на себя. Благодаря им цилиндры и гидравлические шланги полностью защищены от механического повреждения.
Стальные кожухи для защиты гидроцилиндров
Полная защита цилиндров и гидравлических шлангов предохраняет от несчастных случаев
Передовые разработки для современных решений
В ходе разгрузочно — погрузочных работ могут возникать проблемы, и самое главное — найти им своевременное, а лучше превентивное решение. Компания Negrini оснащает свои грейферные ковши различными предохранительными системами и элементами, которых нет у многих производителей грейферов. Непрерывные исследования по улучшению качества, представленные во всем ассортименте продуктов, предлагаемых Negrini, привели к динамичным, инновационным и передовым разработкам в области машиностроения.
Microsoft Word — CEDA_2006_Clamshell.doc
%PDF-1.7
%
137 0 объект
>/OCGs[1568 0 R]>>/StructTreeRoot 1150 0 R/Тип/Каталог/PageLabels 125 0 R>>
эндообъект
139 0 объект
>
эндообъект
134 0 объект
>поток
Acrobat Distiller 6.0 (Windows)2006-08-30T22:03:02+02:00PScript5.dll Версия 5.2.22010-04-12T13:43:55+02:002010-04-12T13:43:55+02:00uuid: 1c25c3aa-9595-4076-aaf7-21fef911d24euuid:e41bee6e-02df-4bb9-b314-957f6b2b73b3application/pdf
конечный поток
эндообъект
1571 0 объект
>/Кодировка>>>>>
эндообъект
127 0 объект
>
эндообъект
1150 0 объект
>
эндообъект
125 0 объект
>
эндообъект
126 0 объект
>
эндообъект
1151 0 объект
>
эндообъект
1551 0 объект
>
эндообъект
1562 0 объект
>
эндообъект
1563 0 объект
>
эндообъект
1567 0 объект
>
эндообъект
1564 0 объект
>
эндообъект
1565 0 объект
>
эндообъект
1566 0 объект
>
эндообъект
1552 0 объект
>
эндообъект
1554 0 объект
>
эндообъект
1556 0 объект
>
эндообъект
1558 0 объект
>
эндообъект
1560 0 объект
>
эндообъект
1561 0 объект
>
эндообъект
1446 0 объект
>>]/P 1444 0 R/S/Link/Pg 747 0 R>>
эндообъект
1437 0 объект
>>]/P 1435 0 R/S/Link/Pg 747 0 R>>
эндообъект
1440 0 объект
>>]/P 1438 0 R/S/Link/Pg 747 0 R>>
эндообъект
1443 0 объект
>>]/P 1441 0 R/S/Link/Pg 747 0 R>>
эндообъект
1428 0 объект
>]/P 1426 0 R/S/Link/Pg 747 0 R>>
эндообъект
1431 0 объект
>>]/стр. 14290 R/S/Link/Pg 747 0 R>>
эндообъект
1434 0 объект
>>]/P 1432 0 R/S/Link/Pg 747 0 R>>
эндообъект
1453 0 объект
>>]/P 1451 0 R/S/Link/Pg 747 0 R>>
эндообъект
1471 0 объект
>>]/P 1469 0 R/S/Link/Pg 747 0 R>>
эндообъект
1474 0 объект
>>]/P 1472 0 R/S/Link/Pg 747 0 R>>
эндообъект
1462 0 объект
>>]/P 1460 0 R/S/Link/Pg 747 0 R>>
эндообъект
1465 0 объект
>>]/P 1463 0 R/S/Link/Pg 747 0 R>>
эндообъект
1468 0 объект
>>]/P 1466 0 R/S/Link/Pg 747 0 R>>
эндообъект
1477 0 объект
>>]/P 1475 0 R/S/Link/Pg 747 0 R>>
эндообъект
1522 0 объект
>]/P 1520 0 R/S/Link/Pg 187 0 R>>
эндообъект
1516 0 объект
>>]/P 1514 0 R/S/Link/Pg 187 0 R>>
эндообъект
1507 0 объект
>>]/P 1505 0 R/S/Link/Pg 187 0 R>>
эндообъект
15190 объект
>>]/P 1517 0 R/S/Link/Pg 187 0 R>>
эндообъект
1510 0 объект
>]/P 1508 0 R/S/Link/Pg 187 0 R>>
эндообъект
1513 0 объект
>]/P 1511 0 R/S/Link/Pg 187 0 R>>
эндообъект
1504 0 объект
>>]/P 1502 0 R/S/Link/Pg 187 0 R>>
эндообъект
1501 0 объект
>>>]/P 1499 0 R/S/Link/Pg 187 0 R>>
эндообъект
1498 0 объект
>>]/P 1496 0 R/S/Link/Pg 187 0 R>>
эндообъект
1495 0 объект
>>]/P 1493 0 R/S/Link/Pg 187 0 R>>
эндообъект
1490 0 объект
>>]/P 1488 0 R/S/Link/Pg 187 0 R>>
эндообъект
1480 0 объект
>]/P 1478 0 R/S/Link/Pg 187 0 R>>
эндообъект
1487 0 объект
>>]/P 1485 0 R/S/Link/Pg 187 0 R>>
эндообъект
1525 0 объект
>>]/P 1523 0 R/S/Link/Pg 158 0 R>>
эндообъект
1528 0 объект
>]/P 1526 0 R/S/Link/Pg 158 0 R>>
эндообъект
1531 0 объект
>>]/P 1529 0 R/S/Link/Pg 158 0 R>>
эндообъект
1534 0 объект
>>]/P 1532 0 R/S/Link/Pg 158 0 R>>
эндообъект
12490 объект
>>]/P 1247 0 R/S/Link/Pg 1034 0 R>>
эндообъект
1226 0 объект
>>]/P 1224 0 R/S/Link/Pg 1034 0 R>>
эндообъект
1253 0 объект
>>]/P 1251 0 R/S/Link/Pg 1034 0 R>>
эндообъект
1257 0 объект
>>]/P 1255 0 R/S/Link/Pg 1034 0 R>>
эндообъект
1258 0 объект
>]/P 1255 0 R/S/Link/Pg 1034 0 R>>
эндообъект
1223 0 объект
>]/P 1220 0 R/S/Link/Pg 1034 0 R>>
эндообъект
1242 0 объект
>]/P 1239 0 R/S/Link/Pg 1034 0 R>>
эндообъект
12390 объект
>
эндообъект
1034 0 объект
>/Шрифт>/ProcSet[/PDF/Text]/ExtGState>>>/Тип/Страница>>
эндообъект
1036 0 объект
[1093 0 Р 1091 0 Р 1089 0 Р 1087 0 Р 1085 0 Р 1083 0 Р 1081 0 Р 1079 0 Р 1077 0 Р 1075 0 Р 1073 0 Р 1071 0 Р 1069 0 Р 1067 0 Р 1065 0 Р 1063 0 0 R 1059 0 R 1057 0 R 1055 0 R 1053 0 R 1051 0 R 1049 0 R 1047 0 R 1045 0 R 1043 0 R 1041 0 R 1039 0 R 1037 0 R]
эндообъект
130 0 объект
>
эндообъект
1913 0 объект
>поток
HWnH}7Âڕ6` ي3Ύc#$$T2cb~BR2uGS:_%jFr!bJwgԅ_Kb~dǁǙ:hQWqل’PۇhW:+cY|L7 />DQPq`8 Ƀ:mωv:Z4ukRКПП’^TnJʷ)M;
uzV-VP/ELJQ ^&Jkrʠ44)1&wۿ!ɬַ̚M|i»dBmf»y(*NEVDŽQ CkQل%˺dJwK_XxcwE9=zN`S쩛yVȷi۫551UZU]6 ̃ЛķMYKQ(w,sr̥fӻhX7#»wC4bᵺN)*yIwH ά{?Cnqr9,Tڃ. x4فyvP_WL|RHcfZEO_ٲD·Fs@Pu&[@Caqvv Ӥ,&2>0gH=;2 9000gH=;2 9000gH=;2 миллисекундные движения моллюсков в лиганд-связывающих доменах ионотропных рецепторов глутамата
Abstract
Ионотропные рецепторы глутамата (iGluRs) опосредуют передачу сигнала в головном мозге и являются важными мишенями для лекарств Структурные исследования показывают снимки iGluRs, которые обеспечивают механистическое понимание стробирования, но быстрые движения, управляющие механизмом рецептора, в значительной степени неуловимы.Здесь мы обнаруживаем кинетику конформационных изменений изолированных лигандсвязывающих доменов в форме моллюска (LBD) из трех основных подтипов iGluR, которые инициируют стробирование при связывании агонистов.Мы разрабатываем флуоресценцию зонды для измерения движения доменов с помощью наносекундной флуоресцентной корреляционной спектроскопии Мы наблюдаем широкий кинетический спектр динамики LBD, которая лежит в основе активации iGluRs. ции и замораживания при связывании полных и частичных агонистов. Мы раскрываем аллостерическое связывание в гетеродимерах NMDA LBD, где связывание L-глутамата с LBD GluN2A останавливает движение моллюсков глицин-связывающего LBD GluN1. Наши результаты показывают быструю динамику LBD в iGluR и предполагают механизм отрицательной аллостерической кооперативности в рецепторах NMDA.
Введение
Передача сигнала в возбуждающих синапсах опосредована ионотропными рецепторами глутамата (iGluRs), которые повсеместно экспрессируются в центральной нервной системе 1,2 . iGluR представляют собой управляемые лигандом ионные каналы, которые играют ключевую роль в развитии мозга и когнитивных функциях более высокого порядка, включая обучение и память. Неисправность рецепторов способствует различным расстройствам головного мозга, таким как эпилепсия, инсульт, болезнь Альцгеймера и шизофрения 3 . Следовательно, iGluRs являются важными мишенями для разработки лекарств. Основываясь на фармакологии и структурной гомологии, iGluRs делятся на три основных подтипа, а именно рецепторы AMPA, kainate и NMDA, все из которых образуют трансмембранные тетрамерные сборки 1 . Каждая субъединица рецептора построена из полуавтономных доменов, соединенных гибкими линкерами. Одна субъединица состоит из внеклеточного N-концевого и лиганд-связывающего доменов, трансмембранного ионного канала и внутриклеточного С-концевого домена (NTD, LBD, TMD и CTD соответственно) 4 . Внеклеточные NTD и LBD представляют собой димерные двухлопастные структуры, напоминающие раковины моллюсков. Двухлопастная форма LBD структурно сходна с бактериальными периплазматическими белками, связывающими аминокислоты 5 . Два слоя внеклеточных доменов iGluR, образованные NTD и LBD соответственно, организованы как димеры димеров. Активация рецептора запускается связыванием агониста с междолевой щелью LBD. Закрытие грейфера переносится на ворота за счет вращения нижней доли D2 9 вверх.0017 6 , запускающий поток ионов для деполяризации постсинаптической клетки, обеспечивающий передачу сигнала 7 .
За последние два десятилетия структурные исследования, включающие рентгеновскую кристаллографию и криоэлектронную микроскопию (крио-ЭМ) вместе с электрофизиологией, позволили получить детальное представление о взаимосвязи структура-функция iGluRs 4,8 . Кристаллическая и крио-ЭМ структуры гомо- и гетеромерных iGluR из AMPA 9,10,11,12,13,14 , NMDA 15,16,17,18,19 и каинат 20 подтипов представили механизмы управления воротами. iGluR-опосредованная передача сигнала уходит своими корнями в сложную сеть конформационных движений отдельных доменов, которые ускользают от экспериментального наблюдения. Начинают определяться динамика отдельных доменов, их междоменная связь и модуляция путем связывания агонистов и антагонистов 21 . Исследования резонансного переноса энергии флуоресценции одиночной молекулы (smFRET) предоставили дополнительную информацию о переходах между структурными конечными состояниями в растворе и выявили их конформационную гетерогенность 22 .
Особое внимание уделяется LBD, «мышце» рецептора, с движениями домена, обеспечивающими триггер, который инициирует открытие каналов 23 . Множество кристаллических структур изолированных LBD в комплексе с различными ант/агонистами показывают, что домен может принимать различные конформации 3,24 . Однако степень их заполнения в растворе и временные шкалы их взаимного превращения остаются неясными. Моделирование молекулярной динамики предполагает, что LBD заполняет более расширенный ансамбль конформаций, чем наблюдается экспериментально 25 .
Кинетика гейтирования различается между подтипами iGluR 8 и зависит от природы агониста внутри подтипа 26 . AMPA/каинатные рецепторы деактивируются в течение нескольких мс, в то время как для рецепторов NMDA требуются сотни мс 2 . Быстрая кинетика активации рецепторов 1 позволяет предположить, что элементарная динамика моллюсков LBD, лежащая в основе стробирования, имеет столь же быструю временную шкалу мс или субмс. Структурные исследования предполагают, что амплитуды движений лепестков LBD составляют порядка одного нанометра (нм) 1,27,28 . Это требует спектроскопических методов с высоким разрешением, чувствительных во временном масштабе до миллисекунд и пространственном масштабе в одну нанометр, для обнаружения функционально значимых движений лепестков.
Здесь мы разработали флуоресцентные зонды, которые в сочетании с флуоресцентной корреляционной спектроскопией (FCS) отвечают этим требованиям к высокому разрешению. Мы использовали чувствительный к окружающей среде оксазиновый флуорофор, который мы пометили специфично для устьев изолированных моллюсков LBD из трех основных подтипов iGluR, а именно AMPA, каината и NMDA. Метка преобразует конформационные флуктуации LBD в флуктуации флуоресценции, которые обнаруживаются с помощью FCS 9.0017 29 . Мы описываем выраженные субмиллисекундные флуктуации в апо-состоянии LBD всех трех подтипов и изучаем их модуляцию при связывании и димеризации агонистов. Мы также раскрываем путь аллостерической коммуникации в динамике NMDA LBD через интерфейс димеризации.
Результаты
Субмиллисекундные движения моллюсков LBD iGluR
Для обнаружения межлепестковых движений между верхними долями D1 и нижними долями D2 (конформационная динамика моллюска) мы использовали экологически чувствительный флуорофор AttoOxa11 (Atto-Tec). который сообщает об изменениях в своем микроокружении изменением интенсивности флуоресцентного излучения в дальнем красном диапазоне спектра. Флуоресценция AttoOxa11 эффективно гасится при образовании ван-дер-ваальсового контакта с боковой цепью триптофана (Trp) посредством фотоиндуцированного переноса электронов (ПЭТ) (рис. 1а) 30 . Быстрые конформационные изменения, которые сопровождаются образованием и нарушением взаимодействия флуорофора/Trp, приводят к флуктуациям флуоресценции ПЭТ, которые можно обнаружить с помощью FCS (PET-FCS) 29,31 . Мы ввели AttoOxa11 в долю D1 подтипов iGluR LBD путем мутации боковой цепи, подверженной воздействию растворителя, на цистеин (Cys) и модификации ее тиол-реактивным AttoOxa11. Природный тушитель Trp (W) был введен в долю D2 (рис. 1b–d). Сайты маркировки в нашем дизайне репортера напоминают структурные координаты, выбранные в качестве меры динамики моллюсков GluA2 LBD в предыдущих компьютерных симуляциях молекулярной динамики 25 . Нативные боковые цепи Cys в LBD либо скрыты, либо образуют структурные дисульфиды и, таким образом, не мешают сайт-специфической модификации сконструированного Cys. Это было очевидно из контрольных экспериментов, в которых испытания модификации LBD дикого типа давали только ~ 10% меченого белка, тогда как LBD, содержащие сконструированные остатки Cys, давали ~ 60% меченого белка (дополнительная таблица 1).
Рис. 1. Конструкция флуоресцентного репортера для обнаружения движений моллюсков LBD iGluR.
a Структурная архитектура iGluRs и расширенный обзор LBD, включая дизайн флуоресцентного репортера PET для движений ракушки. В двухлопастной структуре моллюска LBD сконструированный Cys (красные палочки), модифицированный флуорофором (оранжевая сфера), и сконструированный Trp (синие палочки) в устье раковины моллюска обнаруживают конформационную динамику (серая стрелка) посредством контактно-индуцированного тушение флуоресценции. Связанный агонист показан в виде желтых сфер, а верхняя доля D1 и нижняя доля D2 обозначены. b – d Кристаллические структуры связанных с агонистом GluA2 ( b ), GluK1 ( c ) и GluN1 (d ) LBD в комическом представлении (идентификаторы PDB: 2UXA, 1TXF и 1PB7; для GluA2, GluK1 и GluN1 соответственно). Конструкции ориентированы, как показано на правой стороне панели ( и ). Сайты для модификации флуоресценции (инженерный Cys) показаны красными палочками. Сконструированные остатки Trp показаны синими палочками. Связанный агонист выделен желтыми сферами.
Полноразмерное изображение
Мы провели эксперименты FCS, используя установку конфокального флуоресцентного микроскопа. Для LBD GluA2 и GluK1 AMPA- и каинатного типа одноточечные (т.е. отрицательный контроль) мутанты G446C и K503C, модифицированные AttoOxa11, дали один спад автокорреляционной функции (ACF). Одиночные затухания в АКФ происходили в миллисекундном (мс) временном масштабе и возникали из-за флуктуаций, вызванных броуновской диффузией LBD через фокус детектирования. В то время как ACF от одноточечных мутантов не демонстрировали дополнительных колебаний флуоресценции на шкале времени субмс, двойные мутанты GluA2 G446C-T685W и GluK1 K503C-K734W показали выраженные релаксации субмс значительной амплитуды (рис. 2a, b). Эти дополнительные релаксации возникают из-за движений между лепестками D1 и D2, которые приводят к быстрому формированию и разрушению ван-дер-ваальсовых контактов между AttoOxa11 и сконструированными боковыми цепями W685 и W734. Однако для LBD GluN1 NMDAR флуоресцентно модифицированный одноточечный мутант A480C показал субмс-распад в ACF даже без сконструированного остатка Trp в нижней доле D2 (рис. 2c). Тестирование различных природных аминокислот в качестве потенциальных гасителей флуоресценции показало, что оксазиновые флуорофоры в значительной степени тушатся Trp, немного тушатся тирозином (Tyr, Y), но не тушатся фенилаланином (Phe, F) 32 . В структуре LBD GluN1 мы обнаружили W498/W731 и Y703/Y711 вблизи положения маркировки A480C, остатки, которые могут подавлять флуоресценцию AttoOxa11 (дополнительная рис. 1a). Мы мутировали эти нативные остатки Trp и Tyr по отдельности в фенилаланин (Phe, F), чтобы проверить их потенциальную роль в тушении флуоресценции AttoOxa11 в положении A480C в экспериментах с FCS. Мутация Trp на Phe является структурно консервативной, поскольку она заменяет ароматический индольный фрагмент на ароматический бензол и устраняет потенциальное тушение с помощью Trp. Мы обнаружили, что релаксации sub-ms все еще присутствовали в ACF мутантов F и почти не отличались от релаксаций, обнаруженных для мутанта псевдодикого типа A480C. (Дополнительный рис. 1b, дополнительная таблица 2). Результаты показали, что колебания, обнаруженные у GluN1-A480C, являются результатом изменений в микроокружении чувствительной к окружающей среде метки, таких как полярность, которые были опосредованы конформационными движениями LBD.
Рис. 2: Динамика раскладушки AMPA, каината и NMDA iGluR LBD в апо-состоянии, измеренная с использованием FCS.
а – в АКФ (G(τ)), нормированные на среднее число молекул в фокусе детектирования, зарегистрированные для GluA2 ( a ), GluK1 ( b ) и GluN1 ( c ) LBD. ACF GluA2-LBD-G446C ( a ) и GluK1-LBD-K503C ( b ) показаны синим цветом. ACF GluA2-LBD-G446C-T685W ( a ) и GluK1-LBD-K503C-K734W ( b ) показаны красным цветом. ACF GluN1-A480C показан красным ( c ). Черные линии соответствуют данным с использованием модели молекулярной диффузии, содержащей сумму от одной до четырех одноэкспоненциальных релаксаций (указанных в ( d – f )). d – f Амплитуды (заштрихованные столбцы) и соответствующие константы времени (незакрашенные столбцы), полученные в результате подгонки обнаруженных экспоненциальных фаз в АКФ GluA2 ( d ), GluK1 ( e ) и GluN1 ( ф ) LBD. Наборы данных и цветовой код соответствуют ACF, показанным на панелях ( a – c ). Крестики (X) обозначают отсутствующую (не обнаруженную) кинетику. Столбики погрешностей s.d. трех измерений ( n = 3).
Полноразмерное изображение
Релаксации субмс, очевидные в АКФ GluA2 и LBD GluN1, хорошо описывались суммой трех одноэкспоненциальных распадов (рис. 2a, c, d, f). Для описания релаксаций LBD GluK1 потребовалась сумма четырех одноэкспоненциальных распадов (рис. 2b, e). Уменьшение количества экспонент в применяемой модели подгонки приводило к отклонениям подгонки от данных, а добавление дополнительной экспоненты приводило либо к отсутствию значимого улучшения, либо к переподгонке, что проявлялось в появлении физически необоснованных параметров подгонки ( Дополнительный рис. 2). Следовательно, мы сочли трехэкспоненциальную модель при описании данных LBD GluA2 и GluN1, а четырехэкспоненциальную модель при описании данных LBD GluK1 наиболее подходящей. В целом, мы обнаружили, что режимы движения раковины в этом семействе LBD были во временных масштабах ~100 мкс, ~10 мкс и ~1 мкс. Мы наблюдали специфические для подтипа вариации постоянных времени и амплитуд (рис. 2d – f).
Связывание полных и частичных агонистов останавливает движение моллюска
Затем мы исследовали влияние связывания лиганда на динамику моллюска LBD. С этой целью мы применяли насыщающие концентрации агониста к флуоресцентно модифицированным LBD и регистрировали ACF с использованием FCS. l-глутамат (Glu) применяли в качестве агониста для AMPA- и каинатных LBD (конструкции GluA2-G446C-T685W и GluK1-K503C-K734W), а глицин (Gly) наносили на NMDA GluN1-A480C LBD. Glu и Gly не влияли напрямую на флуоресценцию метки AttoOxa11, что было очевидно из контрольных экспериментов FCS, в которых мы применяли агонисты к флуоресцентно модифицированным конструкциям, не содержащим сконструированного Trp, или только к красителю (дополнительная рис. 3). Для Trp-содержащих мутантов мы наблюдали исчезновение суб-мс-распадов ACF при связывании агониста с LBD, что контрастировало с состоянием апо, которое демонстрировало мкс-релаксации значительной амплитуды (рис. 3). Каинат GluK1 LBD продемонстрировал потерю мкс релаксации при связывании Glu, подобно другим гомологам. Но, в отличие от гомологов, увеличилась амплитуда 200-нс кинетической фазы (рис. 3б, д). Это указывало на существенную подвижность в связанном с агонистом состоянии предположительно локального структурного элемента GluK1 LBD, которая колебалась во временной шкале субмкс. Связанные с агонистом LBD GluA2 и GluN1 демонстрировали остаточные флуктуации малых амплитуд с соответствующими постоянными времени на временной шкале 300 нс и 5 мкс (рис. 3d, f).
Рис. 3: Влияние связывания агониста на динамику LBD iGluR.
а – в АКФ, (G(τ)), нормированные на среднее число молекул в фокусе детектирования, записанные по GluA2-LBD-G446C-T685W ( a ), GluK1-LBD-K503C -K734W ( b ) и GluN1-A480C ( c ) LBD в отсутствие (красный) и в присутствии (голубой) агониста. LBD GluA2 и GluK1 имели Glu в качестве агониста. GluN1 LBD содержал Gly в качестве агониста. Черные линии соответствуют данным с использованием модели молекулярной диффузии, содержащей сумму от одной до четырех одноэкспоненциальных релаксаций (указанных на ( д – ф )). d – f Амплитуды (черные столбцы) и соответствующие постоянные времени (светлые столбцы) экспоненциальных распадов АКФ GluA2 ( d ), GluK1 ( e ) и GluN1 ( f ) LBD. Наборы данных и цветовой код соответствуют ACF, показанным на панелях ( a – c ). Крестики (X) обозначают отсутствующую (не обнаруженную) кинетику. Столбики погрешностей s.d. трех измерений ( n = 3). г Нормализованные ACF, зарегистрированные для GluA2-LBD-G446C-T685W в отсутствие (красный) и в присутствии квисквалата (синий), Glu (голубой), виллардиина (пурпурный), 5-йодвиллардина (зеленый) и каината ( апельсин). Черные линии соответствуют данным с использованием модели молекулярной диффузии, содержащей сумму от одной до трех одноэкспоненциальных релаксаций.
Изображение в полный размер
Чтобы исключить артефакты, вызванные зондом, и проверить функциональность модифицированных LBD, мы провели эксперименты по титрованию лиганда, в которых мы добавили возрастающие концентрации агониста к флуоресцентно модифицированным конструкциям. Мы использовали величину субмс амплитуд флуктуаций в качестве меры для связанного/несвязанного состояния. Постепенные потери суб-мс амплитуд с увеличением концентрации агонистов были хорошо описаны моделью изотермы связывания (дополнительная рис. 4). Анализы дали равновесные константы диссоциации ( K d ) для связывания агонистов, которые находились в диапазоне значений, указанных в литературе. То есть мы обнаружили K D (GLUA2-LBD-G446C-T685W/GLU) = 2,4 ± 0,7 мкм (литература 33,34,35,36,37 : K D = 0,18: K D = 0,18: K D =. мкм), K D (Glun1-A480C/GLY) = 13 ± 3 мкм (литература 38 : K D = 26 мкм) и K 99999999999999999 гг. K503C-K734W/Glu) = 7 ± 3 мкМ. Незначительные расхождения между нашими значениями и значениями из литературы могут быть объяснены различными условиями раствора, применяемыми в разных экспериментальных условиях, или модификацией флуоресценции. Мы пришли к выводу, что исследованные домены после модификации оставались полностью функциональными.
В совокупности наши результаты показали, что внутри этих гомологов iGluR LBD конформационные движения µs высокодинамичных апо-состояний были потеряны при связывании агонистов. Это проявлялось либо в полной потере кинетических фаз, либо в ослабленных экспоненциальных сигналах уменьшенных амплитуд затухания.
Затем мы сравнили влияние связывания полных и частичных агонистов 1 на динамику моллюсков GluA2 LBD. Мы применили насыщающие концентрации агонистов квисквалата, l-Glu, виллардиина, 5-йодвиллардиина и каината (перечислены в порядке убывания эффективности) к образцам GluA2-G446C-T685W и записали ACF с использованием FCS. Для всех пяти агонистов мы наблюдали аналогичное замедление движений моллюсков в мкс (рис. 3g, таблица 1). Кинетика остаточных микросекундных флуктуаций LBD GluA2 в присутствии квисквалата и 5-иодовиллардиина имела амплитуды <10%, что мы считаем слишком малым для надежного определения движения. Таким образом, мы не обнаружили заметных различий в способах действия частичных агонистов по сравнению с полными агонистами в отношении модуляции конформационной динамики LBD.
Таблица 1. Кинетика субмсекционных движений моллюска GluA2-LBD-G446C-T685W в присутствии частичных и полных агонистов.
Полноразмерный стол
Димеризация замедляет движения моллюсков LBD
Внутри полноразмерных сборок iGluR LBD образуют димеры 9 . Мы исследовали влияние димеризации LBD на динамику их ракушечника. Изолированные LBD из подтипов AMPA и NMDA iGluR не образуют димеров в растворе вплоть до концентрации белка в мг/мл 39,40 . Мутация L483Y на поверхности димера GluA2 LBD стабилизирует гомодимер в растворе ( K d = 0,03 мкМ) 39 . Сходным образом, мутации N521Y и E516Y в GluN1 и GluN2A LBD вызывают образование специфических гетеродимеров, т.е. образование сборки GluN1-GluN2A LBD 40 . Мы ввели мутации L483Y, N521Y и E516Y в LBD GluA2, GluN1 и GluN2A соответственно. Динамику гомодимеров, образованных LBD GluA2, исследовали путем регистрации ACF GluA2-LBD-G446C-T685W-L483Y в присутствии избытка GluA2-LBD-L483Y (рис. 4а). Динамику гетеродимеров, образованных LBD GluN1/GluN2A, исследовали путем измерения ACF GluN1-LBD-A480C-N521Y в присутствии избытка GluN2A-LBD-E516Y (рис. 4b). Постоянная времени диффузии, τ D , глобулы прямо пропорциональны ее гидродинамическому радиусу, R h 41 . Мы определили значения R h для Glu-связанного мономера и димера GluA2-LBD, а также для Gly-связанного мономера GluN1-LBD и для Gly/Glu-связанного гетеродимера GluN1/GluN2A LBD. Мы сравнили эти значения с рассчитанными по кристаллическим структурам (таблица 2). R h LBD GluA2, измеренная в растворе с использованием FCS, была значительно больше, чем значение, рассчитанное по структуре. Этот вывод согласуется с ансамблем динамических и более расширенных конформаций LBD в растворе, чем в кристалле. Этот вывод был также предсказан моделированием молекулярной динамики 25 . Для димеров LBD мы наблюдали увеличение R h на ~ 30% по сравнению с мономерами (таблица 2). Это было ожидаемо, потому что удвоение молекулярной массы глобулы приводит к \(\root 3 \of {2}\)-кратному (26%) увеличению R h 41 .
Рис. 4: Динамика раскладушки димерных LBD iGluR.
a АКФ, G(τ), нормированные на среднее число молекул в фокусе детекции, зарегистрированные для GluA2-LBD-G446C-T685W-L483Y (апомономер, красный) и в присутствии избытка GluA2-LBD- L483Y (апо-димер, оранжевый). ACF димера в присутствии агонистов Glu и AMPA показаны голубым и серым цветом (димеры, связанные с агонистом). b ACF записаны для GluN1-LBD-A480C-N521Y (апо-мономер, красный) и в присутствии избытка GluN2A-LBD-E516Y (апо-гетеро-димер, оранжевый). АКФ гетеродимера GluN1-GluN2A в присутствии агониста Gly показан голубым цветом. Черные линии соответствуют данным с использованием модели молекулярной диффузии, содержащей сумму от одного до трех одноэкспоненциальных распадов (указанных в ( c , d )). c Амплитуды (закрашенные столбцы) и соответствующие константы времени (незакрашенные столбцы) экспоненциального распада АКФ мономерного и димерного LBD GluA2 в связанных состояниях апо и агониста. Наборы данных и цветовой код соответствуют ACF, показанным на панели ( и ). Крестики (X) обозначают отсутствующую (не обнаруженную) кинетику. d Амплитуды (закрашенные столбцы) и соответствующие константы времени (незакрашенные столбцы) экспоненциального распада АКФ мономерного и гетеродимерного GluN1 LBD в состояниях, связанных с апо и агонистом. Наборы данных и цветовой код соответствуют ACF, показанным на панели ( b ). Крестики (X) обозначают отсутствующую (не обнаруженную) кинетику. Столбики погрешностей s.d. трех измерений ( n = 3).
Полноразмерное изображение
Таблица 2. Гидродинамические радиусы LBD AMPA и NMDA, измеренные с помощью FCS и сравненные со значениями, рассчитанными на основе доступных рентгеновских структур.
Полноразмерная таблица
Мы обнаружили, что картина суб-мс кинетики движений моллюсков LBD, обнаруженная в апомономерах, сохранялась в димерах (рис. 4c, d). Однако конформационные движения замедлялись, о чем свидетельствовало увеличение постоянных времени всех трех суб-мс-релаксаций при димеризации. Соответствующие амплитуды уменьшались при димеризации (рис. 4в, г). Связывание агониста останавливало конформационную динамику µs в димерах, подобно тому, что мы наблюдали для мономеров, о чем свидетельствует исчезновение релаксаций µs (рис. 4). Мы также применяли более эффективный агонист AMPA 9.0017 26 к GluA2 LBD. Мы обнаружили, что изменения динамики были практически неотличимы от тех, которые наблюдались при связывании Glu: мы наблюдали остановку конформационных релаксаций двух микросекунд. Остаточные наносекундные флуктуации, которые имели аналогичные амплитуды и постоянные времени (рис. 4c).
Аллостерическая передача движений моллюсков в LBD NMDA
LBD NMDA образуют облигатные гетеродимеры, в отличие от AMPA- и каинатных подтипов, которые образуют как гомо-, так и гетеродимеры. В NMDAR LBD GluN1 связывает глицин (Gly), тогда как LBD GluN2A связывает l-глутамат (Glu), а изолированные LBD GluN1 и GluN2A могут собираться в гетеродимеры в растворе 9.0017 40 (рис. 4). Это дало возможность изучить аллостерическую связь между двумя LBD, то есть влияние, которое Glu-связывание с LBD GluN2A оказывает на динамику LBD GluN1 (рис. 5a). Мы обнаружили, что Gly, полный агонист GluN1, как и ожидалось, останавливает движения моллюсков LBD GluN1 внутри димера GluN1/GluN2A. Интересно, однако, что связывание Glu с GluN2A в гетеродимере остановило движение моллюска апо GluN1 LBD внутри димера GluN1/GluN2A (рис. 5b, c). Наблюдение выявило междимерную аллостерию. Аллостерический эффект проявляется в потере ~100 мкс и ~10 мкс кинетических фаз LBD апо-GluN1 при связывании Glu с LBD GluN2A внутри димера, аналогично наблюдаемому для связывания Gly с LBD GluN1. (рис. 5с). В контрольном эксперименте мы нанесли Glu на мономер GluN1-LBD-A480C-N521Y и не обнаружили никакого влияния на движения раскладушки (рис. 5d). Результат подтвердил, что остановка движения GluN1, вызванная связыванием Glu с LBD GluN2A, действительно была аллостерическим эффектом и не индуцировалась прямым взаимодействием Glu с LBD GluN1.
Рис. 5: Аллостерическая связь динамики раскладушки в димерах NMDA iGluR LBD.
a Дизайн репортерной системы, которая определяет влияние связывания агониста с LBD GluN2A на динамику LBD GluN1. Метка флуоресценции на LBD GluN1 показана красной сферой, а динамика раскладушки LBD — красной стрелкой. Связывание агониста Glu показано желтой стрелкой. b ACF, G(τ), нормализованное к среднему количеству молекул в фокусе детекции, записанное из LBD GluN1 внутри димера GluN1/GluN2A (оранжевые данные, GluN1-LBD-A480C-N521Y/GluN2A-LBD-E516Y ). ACF гетеродимера, измеренные в присутствии агониста Glu или Gly, показаны синим и голубым цветом соответственно. Черные линии соответствуют данным с использованием модели молекулярной диффузии, содержащей сумму от одного до трех одноэкспоненциальных распадов (указаны на панели ( с )). c Амплитуды (темные столбцы) и соответствующие постоянные времени (незакрашенные столбцы) экспоненциальных спадов АКФ, записанных для димера GluN1/GluN2A. Наборы данных и цветовой код соответствуют ACF, показанным на панели ( b ). Крестики (X) обозначают отсутствующую (не обнаруженную) кинетику. d ACF мономерного апо GluN1-LBD (конструкция A480C-N521Y), измеренные в отсутствие (красный) и в присутствии (синий) 1 мМ Glu. e ACF димера GluN1/GluN2A LBD, содержащего мутацию Y535S (конструкция GluN1-LBD-A480C-N521Y-Y535S/GluN2A-LBD-E516Y), измеренная в апо-состоянии (оранжевый), в присутствии Glu (синий) или при присутствии Gly (голубого). Черные линии соответствуют данным с использованием модели молекулярной диффузии, содержащей сумму двух или трех одноэкспоненциальных распадов (указаны на панели ( ф )). f Амплитуды (закрашенные столбцы) и соответствующие константы времени (незакрашенные столбцы) экспоненциального распада ACF, записанные для димера GluN1/GluN2A LBD, содержащего мутацию Y535S. Наборы данных и цветовой код соответствуют ACF, показанным на панели ( e ). Крест (X) обозначает отсутствующую (не обнаруженную) кинетику. Столбики погрешностей s.d. трех измерений ( n = 3).
Изображение с полным размером
Специфический тирозиновый остаток в LBD GluN1, Y535, расположенный на границе димеризации, играет важную роль в модулировании дезактивации рецептора 40 . Чтобы исследовать роль этого остатка в аллостерии динамики между GluN2A и GluN1 LBD, мы создали мутант GluN1, Y535S, который удаляет ароматическую боковую цепь 40 (конструкция GluN1-LBD-A480C-N521Y-Y535S). Мутация Y535S не влияла на динамику мономера в виде раковины моллюска, о чем свидетельствовали умеренные изменения динамики LBD GluN1 и сохранение остановки движений при связывании Gly (дополнительная рис. 5). Кинетика движений моллюсков димеров LBD GluN1/GluN2A, содержащих мутацию Y535S (GluN1-Y535S/GluN2A), также сохранялась (ср. рис. 5e, f и рис. 4b, d), показывая, что эта мутация не нарушала динамику димер. Более того, применение Gly к димеру GluN1-Y535S/GluN2A остановило кинетические фазы движения моллюска GluN1 длительностью ~100 мкс и ~10 мкс (рис. 5e, f). Это был тот же эффект, что и для димера GluN1/GluN2A без мутации Y535S (рис. 5b, c). Интересно, однако, что применение Glu к GluN1-Y535S/GluN2A практически не влияло на движения моллюсков GluN1 (рис. 5e, f), что отличалось от поведения GluN1/GluN2A без мутации Y535S (рис. 5b, в). Следовательно, делеция ароматической боковой цепи Y535 разъединяет передачу движений моллюска от GluN2A к GluN1 LBD, подчеркивая критическую роль Y535 в аллостерическом связывании NMDAR LBD.
Обсуждение
Множество структурных исследований iGluRs подтверждают модель, в которой связывание агониста с LBD стабилизирует конформацию с закрытой щелью, процесс, который в конечном счете вызывает открытие ворот канала 24 . Структурные исследования показывают, что амплитуда движения лепестков составляет порядка одного нанометра 28 . Быстрая миллисекундная кинетика 1 активации рецепторов указывает на то, что лежащие в основе движения моллюсков LBD должны иметь аналогичный временной масштаб. Моллюсковые движения изолированных доменов, как можно ожидать, будут даже быстрее, потому что меж- и внутрисубъединичные взаимодействия доменов внутри полноразмерных сборок iGluR будут замедлять динамику. Здесь мы использовали наносекундную FCS в сочетании с чувствительной к окружающей среде оксазиновой меткой для исследования динамики LBD в пространственном масштабе в один нанометр и во временном масштабе субмс. Мы обнаружили ярко выраженную мультиэкспоненциальную кинетику реконфигурации LBD в апо-состоянии, которое имело значительные амплитуды релаксации. Поскольку конформационный переход с двумя состояниями следует моноэкспоненциальному ходу времени 42 , количество экспонент суб-мс, обнаруженных с помощью FCS, можно интерпретировать как количество режимов конформационных изменений. Для LBD подтипов AMPA и NMDA мы обнаружили три режима движения, которые были на временной шкале от нс до мкс, тогда как каинатный LBD показал четыре режима движения. FCS измеряет равновесную кинетику, и одноэкспоненциальное затухание ACF можно интерпретировать как происходящее от одного термически активированного конформационного изменения вдоль определенной координаты реакции. В динамике белков порядок временных масштабов конформационных изменений обычно следует порядку рангов пространственных масштабов, на которых они происходят. Петлевые движения и движение вторичной структуры происходят во временном масштабе от нс до мкс, в то время как коллективные движения доменов происходят во временном масштабе от мкс до мс 43 . Моделирование молекулярной динамики, проведенное на LBD NMDA iGluR, предсказывает широкий спектр движений лопастей 44 : в моделировании были идентифицированы три основных компонента, которые классифицируются как изгибание шарнира, подметание и скручивание. Любопытно, что количество основных компонент, выявленных при моделировании, совпадает с количеством экспонент, обнаруженных в наших экспериментах, что позволяет предположить, что наблюдаются одни и те же процессы. Моделирование также показывает расширенные конформации, отобранные с помощью LBD, за пределы того, что наблюдается в структурных исследованиях 25,44 . Спектроскопия smFRET обнаруживает гетерогенный ансамбль конформаций LBD GluA2, которые взаимопревращаются в масштабе времени ~ 100 мс 45 . Эта временная шкала более чем на три порядка медленнее, чем те, которые мы обнаружили здесь с помощью FCS. Расхождение предполагает, что природа конформационных переходов, обнаруженных smFRET и FCS, различна. Однако smFRET в сочетании с быстрой корреляционной спектроскопией обнаруживает быструю междоменную динамику между двумя верхними долями D1 в димерной сборке LBD из метаботропного рецептора глутамата (mGluR) 46 . Шкала времени 50–100 мкс, обнаруженная для междоменных движений D1-D1, согласуется с временными константами, обнаруженными здесь для внутрисубъединичных движений доли D1-D2 LBD iGluR. Интересно, что быстрая динамика D1-D1 была обнаружена в связанных с лигандом активных и покоящихся состояниях сборок mGluR LBD, что показывает, что связанные с лигандом домены остаются подвижными по отношению друг к другу 46 .
Мы обнаружили, что связывание агонистов останавливало движения моллюсков суб-мс всех трех гомологов iGluR LBD, о чем свидетельствовало исчезновение экспоненциальных спадов в ACF (рис. 3). Быстрые, термически активируемые равновесные флуктуации в состояниях апо, обнаруженные с помощью FCS, предполагают механизм конформационного отбора связывания лиганда. Это также было сделано из результатов предыдущей спектроскопии smFRET субмс, проведенной на NMDA LBD 9.0017 47 . Мы наблюдали остаточные конформационные флуктуации в состояниях LBD, связанных с агонистом (рис. 3, 4). Они происходили в основном в сверхбыстром масштабе времени субмкс. Моделирование молекулярной динамики предсказывает остаточную гибкость в состояниях, связанных с агонистом, AMPA и NMDA LBD 23,25,44 , что согласуется с нашими результатами. ЯМР-исследования обнаруживают доказательства гибкости как в состоянии, связанном с апо, так и в состоянии, связанном с агонистом 48 .
Мы обнаружили субмикросекундные флуктуации выраженной амплитуды в связанном с агонистом состоянии GluK1 LBD. Амплитуда была существенно выше по сравнению с амплитудами соответствующих распадов, наблюдаемых в LBD GluA2 и GluN1. Это открытие можно объяснить тем фактом, что сайт мечения флуорофора является частью расширенного сегмента петли GluK1, который на два остатка длиннее по сравнению с теми же сегментами в GluA2 и GluN1 и, таким образом, может обеспечивать дополнительную гибкость (рис. 1c).
Мы обнаружили, что связывание полных агонистов AMPA и Glu оказывает практически неразличимое влияние на динамику LBD: оба агониста тормозят одни и те же режимы движения LBD GluA2 как в мономерном, так и в димерном состоянии. (рис. 4в). Это наблюдение можно объяснить сходством кристаллических структур LBD GluA2 в комплексе с Glu или AMPA 24 , что предполагает сходные способы действия. Связывание частичных агонистов с iGluR снижает амплитуды электрофизиологических сигналов по сравнению со связыванием полных агонистов, но механизм еще полностью не изучен 49 . Связывание частичных агонистов может приводить к более открытой конформации моллюсков LBD по сравнению со связыванием полных агонистов и, следовательно, может запускать ворота канала с меньшей вероятностью. Этот сценарий предложен структурными исследованиями ряда стерически сложных замещенных виллардиинов, связанных с LBD GluA2, которые постепенно обнаруживают более открытые домены 49 . Эксперименты smFRET, проведенные на LBD GluA2, показывают гетерогенный ансамбль структурных состояний, заселяемых при связывании виллардиинов 9.0017 50 , которые, похоже, имеют разные конформации. Альтернативная модель частичного агонизма предполагает, что более низкая стабильность комплекса LBD/частичный агонист снижает эффективность, несмотря на индукцию полностью закрытой раковины моллюска 51 . Третий сценарий может включать более высокую подвижность LBD, связанного с частичным агонистом, по сравнению с полным агонистом, что может привести к более низкой вероятности срабатывания ворот канала. Здесь мы обнаружили, что связывание частичных агонистов, таких как заменители виллардиинов, с LBD GluA2 дает идентичные динамические характеристики по сравнению со связыванием полных агонистов, таких как квисквалат, AMPA или Glu (рис. 3g, таблица 1). Результат подтверждает механизм, посредством которого различная стабильность или выбранные конформации LBD в комплексе с полными или частичными агонистами отвечают за различную эффективность.
Мы наблюдали, что димеризация LBD приводит к увеличению постоянных времени движений моллюсков и уменьшению соответствующих кинетических амплитуд (рис. 4). Это разумно, потому что интерфейс димеризации покрывает шарнирную область раскладушки. Таким образом, димеризация будет ограничивать подвижность долей D1 и D2 относительно друг друга.
Сборка LBD GluN1 и GluN2A в виде гетеродимеров позволила нам изучить аллостерические эффекты. Мы исследовали влияние связывания Glu с LBD GluN2A на динамику моллюсков LBD GluN1 внутри димера. К нашему удивлению, мы обнаружили, что связывание Glu с LBD GluN2A останавливало движения GluN1, подобно тому, как это делал Gly при непосредственном связывании с GluN1 (рис. 5). Ранее предполагалось, что связывание агониста с одной субъединицей LBD может стабилизировать закрытое состояние соседней субъединицы 9 без лиганда. 0017 52,53 . Именно это мы и наблюдали здесь. Далее мы обнаружили, что мутация Y535S, локализованная в шарнирной области LBD GluN1, отменяет аллостерический эффект: связывание Glu с LBD GluN2A в димере GluN1/GluN2A LBD, где ароматическая боковая цепь Y535 была делетирована, практически не оказывает никакого эффекта. на динамику LBD GluN1 (рис. 5). Боковая цепь Y535 в интерфейсе димеризации GluN1/GluN2A заполняет карман, который является мишенью аллостерических модуляторов гомологичного AMPA iGluR LBD 9.0017 40 . Электрофизиология показывает, что мутация Y535S ускоряет дезактивацию NMDA iGluRs и предполагается, что она служит сцеплением между LBD GluN1 и GluN2A 40 . Это предположение согласуется с нашими выводами, показывающими, что Y535 передает индуцированное связыванием лиганда торможение динамики от GluN2A LBD к GluN1 LBD. Исследования smFRET, проведенные на полноразмерных рецепторах, предполагают усиленное конформационное распространение и гибкость LBD GluN1, индуцированные связыванием Glu с GluN2A. Выводы основаны на распространении и уширении гистограмм smFRET, записанных от LBD GluN1 в рецепторе при применении Glu 9.0017 54 . Несоответствие между результатами smFRET и нашими результатами FCS может быть объяснено различными пространственными и временными масштабами, исследованными методами. FRET исследует глобальные конформационные изменения в масштабе 2–10 нм, в то время как контактно-индуцированное тушение активно в масштабе 1 нм 55 . Конформационные состояния, наблюдаемые на гистограммах smFRET, взаимопревращаются медленнее, чем мс, что присуще применяемому методу сбора данных, тогда как FCS обнаруживает быструю динамику на шкале времени нс-мс 31 . Таким образом, вполне вероятно, что разные конформационные подсостояния исследуются разными методами. Более того, поскольку эксперименты с smFRET проводились в контексте полноразмерного рецептора 54 , меж- и внутрисубъединичные взаимодействия меток внутри рецептора могут модулировать интенсивность их флуоресцентного излучения и вносить вклад в гетерогенность гистограмм smFRET. Однако моделирование молекулярной динамики, проведенное в том же исследовании, показывает, что связывание Glu с LBD GluN2A вызывает более закрытую конформацию расщелины LBD GluN1 54 , что согласуется с наблюдаемыми здесь остановленными движениями GluN1 LBD.
Сообщается об отрицательной кооперативности Gly с Glu в NMDA iGluR 2,54,56 . Связывание Glu с GluN2A снижает сродство GluN1 к Gly 40 . Наши результаты предполагают механизм мобильности, лежащий в основе этого явления: связывание Glu с GluN2A LBD останавливает динамику GluN1 LBD через аллостерический путь с участием Y535. В рамках связывания агонистов посредством конформационного отбора сниженная гибкость LBD GluN1 ослабляет его сродство к Gly.
Методы
Мутагенез, синтез и флуоресцентная модификация белков
ДНК-конструкции содержали гены, кодирующие Rattus norvegicus AMPA GluA2, каинат GluK1, NMDA GluN1 и NMDA GluN2A лиганд-связывающие доли S1 и Thr S2, линкер и N-концевой His 6 -tag в составе векторов экспрессии T7 pET22b(+) для конструкции GluA2, GluK1 и GluN1 и pET22b(+)-Sumo для конструкции GluN2A 40 . Одноточечные мутанты были созданы с использованием протокола мутагенеза QuikChange (Stratagene).
LBD и их мутанты были сверхэкспрессированы в клетках Origami 2 (DE) (конструкции AMPA) или Origami B (DE3) (конструкции каината и NMDA) Escherichia coli (Novagen) с использованием системы экспрессии T7. После выращивания бактериальных клеток до OD 600 нм 2,0 в жидкой среде Terrific Broth (TB) (конструкции AMPA) или среде Луриа-Бертани (LB) (конструкции каината и NMDA), содержащей 100 мкг/мл ампициллина (конструкции AMPA) или 50 мкг/мл ампициллина и 12,5 мкг/мл тетрациклина (конструкции каината и NMDA), сверхэкспрессию индуцировали добавлением 0,4 мМ (конструкции AMPA) или 0,5 мМ (конструкции каината и NMDA) изопропил-β-D-1-тиогалактопиранозида (IPTG, Sigma — Олдрич). Затем клетки Origami B инкубировали при 15°C в течение 20 часов, тогда как клетки Origami 2 инкубировали при 18°C в течение 20 часов. После лизиса ультразвуком His 6 -меченый белок выделяли из лизатов бактериальных клеток путем их загрузки на хроматографическую колонку Talon®Superflow™ (Sigma-Aldrich), уравновешенную 20 мМ Tris-HCl, pH 8,0, 300 мМ NaCl, 10 мМ имидазола, содержащего 1 мМ Моногидрат калиевой соли l-глутаминовой кислоты (Sigma-Aldrich) или глицин (Sigma-Aldrich) (конструкции каината и NMDA), или 20 мМ Трис-HCl, pH 8,0, 500 мМ NaCl, 20 мМ имидазол, содержащий L-глутаминовую кислоту моногидрат калиевой соли (конструкции AMPA). В случае каинатных и NMDA-конструкций смолу Talon®Superflow™ промывали, применяя градиент имидазола от 16 мМ до 32 мМ в уравновешивающем буфере. LBD элюировали со смолы, используя 20 мМ Tris-HCl, pH 8,0, 300 мМ NaCl, содержащую 250 мМ имидазола и 1 мМ моногидрат калиевой соли l-глутаминовой кислоты или глицин (конструкции каината и NMDA), или 20 мМ Tris-HCl, pH 8,0, 500 мМ NaCl, содержащий 500 мМ имидазола и 1 мМ моногидрат калиевой соли l-глутаминовой кислоты (конструкции AMPA). Элюат наносили на колонку для эксклюзионной хроматографии (SEC) (HiLoad 26/60 Superdex™ 75) (GE Healthcare) в 20 мМ HEPES, 150 мМ NaCl, pH 7,5, содержащую 1 мМ моногидрат калиевой соли l-глутаминовой кислоты (AMPA). конструкции) или, для каинатных и NMDA-конструкций, подвергают диализу в буфере для анионообменной хроматографии (20 мМ трис-HCl, pH 8,5, 20 мМ NaCl, содержащий 1 мМ моногидрат калиевой соли 1-глутаминовой кислоты или глицин). Во время приготовления конструкций NMDA GluN2A LBD слитый белок сумо удаляли посредством протеолитического расщепления (сумопротеаза Ulp-1). В случае AMPA GluA2, каината GluK1 и NMDA GluN1 LBD, His 6 -tag удаляли протеолитическим расщеплением с использованием тромбина (Sigma-Aldrich). LBD дополнительно очищали с помощью анионообменной хроматографии (5 мл колонка HiTRap™ Q FF, GE Healthcare), применяя градиент от 20 мМ до 500 мМ NaCl в 20 мМ Трис-HCl, рН 8,5, содержащем 1 мМ моногидрат калиевой соли 1-глутаминовой кислоты. или глицин. Объединенные фракции, содержащие LBD, концентрировали с использованием центробежного концентратора с молекулярной массой 10 кДа (Sartorius™ Vivaspin™ 20). Чистоту LBD подтверждали с помощью SDS-PAGE.
Одноточечные мутанты Cys конструкций LBD были модифицированы с использованием тиол-реактивного малеимидного производного красителя AttoOxa11 (Atto-Tec). Мечение проводили в 20 мМ HEPES, 150 мМ NaCl, pH 7,5, содержащем 1 мМ калиевой соли 1-глутаминовой кислоты или глицина и десятикратном молярном избытке восстанавливающего агента трис(2-карбоксиэтил)фосфина (TCEP, Sigma-Aldrich ). Применяли 15-кратный молярный избыток AttoOxa11 по сравнению с LBD. Реакцию мечения проводили в течение 3 ч при 4°С. Избыток красителя удаляли с помощью SEC на колонке Sephadex G-25 (GE Healthcare).
Степень мечения (DOL) LBD определяли по сигналу УФ-поглощения белка и сигналу поглощения видимой области метки конъюгата в соответствии с инструкциями производителя (Atto-Tec):
$${ {{{{{\rm{DOL}}}}}}}}=\frac{c({{{{{{\rm{dye}}}}}}}})}{c({{{{{{{ \rm{белок}}}}}}}})}=\frac{{A}_{{\max}}\cdot {\varepsilon}_{{{{{{{{\rm{prot}}}}}} }}}}{({A}_{280}-{A}_{{\max}}\cdot {{CF}}_{280})\cdot {\varepsilon}_{{\max}}} $$
(1)
где A max – сигнал поглощения конъюгата при максимуме длины волны красителя, ε prot – коэффициент экстинкции белка при 280 нм, A 280 – сигнал поглощения конъюгата при 280 нм, CF 280 — поправочный коэффициент метки, специфичный для красителя, который корректирует вклад красителя, а ε max — коэффициент экстинкции метки в максимуме длины волны.
Эксперименты FCS
Наносекундные FCS проводились на специально изготовленной установке конфокального флуоресцентного микроскопа с применением взаимной корреляции сигналов от двух оптоволоконных лавинных фотодиодных детекторов (Perkin Elmer, SPCM-AQRH-15-FC) для обхода неработающего детектора. -эффекты времени и постимпульса с использованием цифрового аппаратного коррелятора (цифровой реальный коррелятор ALV 5000/60×0 с множеством тау) 57 . Конструкции LBD измеряли в 50 мМ фосфата, рН 7,5, с доведением ионной силы до 200 мМ с использованием хлорида калия. Конструкции LBD GluN1/GluN2A измеряли в 20 мМ HEPES, pH 7,5, с доведением ионной силы до 150 мМ с использованием хлорида натрия. 0,05 % Tween-20 (Sigma-Aldrich) и 0,3 мг/мл альбумина бычьей сыворотки (BSA, Sigma-Aldrich) использовали в качестве буферных добавок для подавления проникновения на поверхность стекла. АКФ регистрировались ок. 1 нМ флуоресцентно модифицированные образцы LBD. Для экспериментов, связанных с димеризацией LBD, к флуоресцентно модифицированной конструкции LBD добавляли избыток обычно 40 мкМ немеченой конструкции LBD. В исследованиях связывания агонистов перед измерением к образцам добавляли 1 мМ агониста. Все измерения проводились при 25 °C. Точный контроль температуры был достигнут с помощью изготовленного на заказ нагревателя объектива. Все образцы были отфильтрованы с помощью шприцевого фильтра с размером пор 0,2 мкм и перед измерениями перенесены на покровное стекло высокой точности. Для каждой экспериментальной установки регистрировали по три АКФ от одного образца ( 9{-1}$$
(2)
N – среднее число молекул в фокусе детектирования, а \({{{{{{\rm{\tau }}}}}}}_{{{ {{{\rm{D}}}}}}}\) представляет постоянную времени диффузии. Применение модели диффузии в двух измерениях имело достаточную точность, поскольку два горизонтальных размера ( x , y ) фокуса детектирования были намного меньше поперечного размера ( х ) в применяемой установке.
АКФ, содержащие дополнительные распады на шкале субмс, были описаны суммой одинарных экспонент с использованием расширенной аналитической модели 9{-1}\left(1+\mathop{\sum}\limits _{{{n}}}{{{a}}}_{{{n}}}\exp \left(-\frac{\ tau }{{\tau }_{n}}\right)\right)$$
(3)
, где a n обозначает амплитуду, а \({{{{{{\rm {\tau }}}}}}}_{{{n}}}\) постоянная времени n -го распада.
В этой модели изменение между флуоресцентной и гасящей флуоресценцию конформацией описывается двухуровневым равновесием, кинетика которого соответствует моноэкспоненциальному затуханию. Амплитуда и постоянная времени экспоненты содержат микроскопические константы скорости включения/выключения флуоресценции. Несколько независимых конформационных релаксаций с двумя состояниями описываются суммой экспонент. Микроскопический анализ констант скорости осложняется тем, что амплитуда распада также модулируется яркостью состояний 31 .
Значения τ D были преобразованы в R h с использованием уравнения Стокса-Эйнштейна 41 . Для калибровки установки мы измерили τ D флуорофора Atto655 в качестве эталона, для которого известен коэффициент диффузии и, следовательно, R h 59 . Для сравнения, радиусы вращения ( R г ) LBD были рассчитаны из доступных кристаллических структур с использованием PyMOL (pymol. org). Значения R h вычислены из R g с использованием теоретического закона подобия для монодисперсной сферы 60 :
$${R}_{h}=\frac{1}{\sqrt{3 /5}}\cdot {R}_{g}$$
(4)
Для определения значений K d из амплитуд релаксации FCS, построенных в зависимости от концентрации агониста, мы применили модель для белка изотерма связывания -лиганда (P + L = PL) 61 :
$$\left[PL\right]=\frac{{[P]}_{t}[L]}{\left[L\right ]+{K}_{d}}$$
(5)
где [PL] — концентрация комплекса белок-лиганд, [P] t — общая концентрация флуоресцентно модифицированного белка, [L] — концентрация лиганда. Изменение амплитуды релаксации FCS 90 147 a 90 148 моделировалось как:
$$\frac{a-{a}_{u}}{{a}_{b}-{a}_{u}}=\ frac{[L]}{\left[L\right]+{K}_{d}}$$
(6)
где a — наблюдаемая амплитуда, a u — амплитуда в несвязанном состоянии и a b – амплитуда в связанном состоянии.
Сводка отчета
Дополнительную информацию о дизайне исследования можно найти в Сводке отчета об исследовании природы, связанной с этой статьей.
Доступность данных
Данные, подтверждающие результаты этого исследования, можно получить у соответствующего автора по обоснованному запросу. Исходные данные фигур доступны в качестве дополнительных данных.
Ссылки
Traynelis, S. F. et al. Ионные каналы глутаматных рецепторов: структура, регуляция и функция. Фарм. 62 , 405–496 (2010).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Erreger, K., Chen, P.E., Wyllie, D.J. & Traynelis, S.F. Управление глутаматными рецепторами. Крит. Преподобный Нейробиол. 16 , 187–224 (2004).
КАС
пабмед
СтатьяGoogle ученый
Грегер И. Х. и Майер М. Л. Структурная биология ионных каналов глутаматных рецепторов: к пониманию механизма. Курс. мнение Структура биол. 57 , 185–195 (2019).
КАС
пабмед
СтатьяGoogle ученый
Stern-Bach, Y. et al. Агонистическая селективность глутаматных рецепторов определяется двумя доменами, структурно родственными бактериальным белкам, связывающим аминокислоты. Нейрон 13 , 1345–1357 (1994).
КАС
пабмед
СтатьяGoogle ученый
Каракас, Э., Риган, М.С. и Фурукава, Х. Новое структурное понимание функции ионотропных рецепторов глутамата. Тренды Биохим. науч. 40 , 328–337 (2015).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Zhu, S. & Gouaux, E. Структура и симметрия определяют принципы работы ионотропных рецепторов глутамата. Нейрофармакология 112 , 11–15 (2017).
КАС
пабмед
СтатьяGoogle ученый
Соболевский А. И., Роскони М. П. и Гуо Э. Рентгеновская структура, симметрия и механизм рецептора глутамата подтипа AMPA. Природа 462 , 745–756 (2009).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Herguedas, B. et al. Структура и организация гетеромерных рецепторов глутамата AMPA-типа. Наука 352 , aad3873 (2016).
ПабМед
ПабМед Центральный
Статья
КАСGoogle ученый
Herguedas, B. et al. Архитектура гетеромерного рецептора GluA1/2 AMPA в комплексе со вспомогательной субъединицей TARP gamma8. Наука 364 , eaav9011 (2019).
Durr, K.L. et al. Структура и динамика АМРА-рецептора GluA2 в состоянии покоя, предоткрытом и десенсибилизированном состояниях. Cell 158 , 778–792 (2014).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Туми, Э. К., Елшанская, М.
В., Грассуччи, Р. А., Франк, Дж. и Соболевский, А. И. Открытие каналов и механизм ворот в рецепторах глутамата подтипа AMPA. Природа 549 , 60–65 (2017).КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Чжан, Д., Уотсон, Дж. Ф., Мэтьюз, П. М., Кэйс, О. и Грегер, И. Х. Гейтирование и модуляция гетерооктамерного рецептора глутамата AMPA. Природа 594 , 454–458 (2021).
КАС
пабмед
СтатьяGoogle ученый
Lee, C.H. et al. Структуры рецепторов NMDA показывают расположение субъединиц и архитектуру пор. Природа 511 , 191–197 (2014).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Zhu, S. et al. Механизм ингибирования и активации NMDA-рецепторов.
Cell 165 , 704–714 (2016).КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Lu, W., Du, J., Goehring, A. & Gouaux, E. Крио-ЭМ структуры тригетеромерного рецептора NMDA и его аллостерическая модуляция. Наука 355 , eaal3729 (2017).
Джалали-Язди Ф., Чоудхури С., Йошиока С. и Гуо Э. Механизмы ингибирования цинком и протонами рецептора GluN1/GluN2A NMDA. Cell 175 , 1520–1532 e1515 (2018).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Каракас Э. и Фурукава Х. Кристаллическая структура ионного канала гетеротетрамерного рецептора NMDA. Наука 344 , 992–997 (2014).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Чоу, Т. Х., Таджима, Н., Ромеро-Эрнандес, А. и Фурукава, Х. Структурная основа функциональных переходов в рецепторах NMDA млекопитающих. Cell 182 , 357–371 e313 (2020).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Маклин, Д. М., Дарем, Р. Дж. и Джаяраман, В. Картирование конформационного ландшафта глутаматных рецепторов с использованием одиночной молекулы FRET. Тренды Неврологии. 42 , 128–139 (2019).
КАС
пабмед
СтатьяGoogle ученый
Освальд, Р. Э.
Гибкость глутамат-связывающего домена. Структура 15 , 1157–1158 (2007).КАС
пабмед
СтатьяGoogle ученый
Армстронг Н. и Гуо Э. Механизмы активации и антагонизма AMPA-чувствительного глутаматного рецептора: кристаллические структуры ядра, связывающего лиганд GluR2. Нейрон 28 , 165–181 (2000).
КАС
пабмед
СтатьяGoogle ученый
Лау, А. Ю. и Ру, Б. Ландшафты свободной энергии, управляющие конформационными изменениями в лиганд-связывающем домене рецептора глутамата. Структура 15 , 1203–1214 (2007).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Zhang, W., Robert, A., Vogensen, S.B. & Howe, J.R. Взаимосвязь между активностью агониста и кинетикой рецептора AMPA.
Биофиз. J. 91 , 1336–1346 (2006).КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Армстронг, Н., Джасти, Дж., Бейх-Франдсен, М. и Гуо, Э. Измерение конформационных изменений, сопровождающих десенсибилизацию в ионотропном рецепторе глутамата. Cell 127 , 85–97 (2006).
КАС
пабмед
СтатьяGoogle ученый
Кумар, Дж. и Майер, М.Л. Функциональные выводы из структур ионных каналов глутаматных рецепторов. Год. Преподобный Физиол. 75 , 313–337 (2013).
КАС
пабмед
СтатьяGoogle ученый
Дженсен, М. Х., Сукумаран, М., Джонсон, К. М., Грегер, И. Х. и Нойвейлер, Х. Внутренние движения в N-концевом домене ионотропного рецептора глутамата, обнаруженные с помощью флуоресцентной корреляционной спектроскопии.
Дж. Мол. биол. 414 , 96–105 (2011).КАС
пабмед
СтатьяGoogle ученый
Дуз, С., Нойвейлер, Х. и Зауэр, М. Тушение флуоресценции фотоиндуцированным переносом электронов: репортер конформационной динамики макромолекул. ChemPhysChem 10 , 1389–1398 (2009).
КАС
пабмед
СтатьяGoogle ученый
Sauer, M. & Neuweiler, H. PET-FCS: исследование быстрых структурных колебаний белков и нуклеиновых кислот путем тушения флуоресценции одной молекулы. Методы Мол. биол. 1076 , 597–615 (2014).
КАС
пабмед
СтатьяGoogle ученый
Марме, Н., Кнемейер, Дж. П., Зауэр, М. и Вольфрум, Дж. Тушение меж- и внутримолекулярной флуоресценции органических красителей триптофаном. Биоконъюгат Хим.
14 , 1133–1139 (2003).КАС
СтатьяGoogle ученый
Abele, R., Keinanen, K. & Madden, D.R. Индуцированная агонистами изомеризация в лиганд-связывающем домене рецептора глутамата. Кинетический и мутагенетический анализ. Дж. Биол. хим. 275 , 21355–21363 (2000).
КАС
пабмед
СтатьяGoogle ученый
Джин, Р., Хорнинг, М., Майер, М.Л. и Гуо, Э. Механизм активации и селективности в лиганд-управляемом ионном канале: структурные и функциональные исследования GluR2 и квасквалата. Биохимия 41 , 15635–15643 (2002).
КАС
пабмед
СтатьяGoogle ученый
Сакакура, М. и др. Структурные механизмы, лежащие в основе изменений активности рецептора глутамата AMPA-типа, вызванных заменами в его лиганд-связывающем домене.
Строение 27 , 1698–1709 e1695 (2019).КАС
пабмед
СтатьяGoogle ученый
Kiyonaka, S. et al. Направленная лигандом химия рецепторов AMPA обеспечивает флуоресцентные биосенсоры живых клеток. ACS Хим. биол. 13 , 1880–1889 (2018).
КАС
пабмед
СтатьяGoogle ученый
Deming, D., Cheng, Q. & Jayaraman, V. Является ли изолированный лиганд-связывающий домен хорошей моделью домена в нативном рецепторе? Дж. Биол. хим. 278 , 17589–17592 (2003 г.).
КАС
пабмед
СтатьяGoogle ученый
Yao, Y. & Mayer, M.L. Характеристика растворимого лиганд-связывающего домена регуляторной субъединицы рецептора NMDA NR3A. J. Neurosci. 26 , 4559–4566 (2006).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Furukawa, H., Singh, S.K., Mancusso, R. & Gouaux, E. Расположение и функция субъединиц в рецепторах NMDA. Природа 438 , 185–192 (2005).
КАС
пабмед
СтатьяGoogle ученый
Teufel, D.P., Johnson, C.M., Lum, J.K. & Neuweiler, H. Вызванный позвоночником коллапс в развернутых белковых цепях. Дж. Мол. биол. 409 , 250–262 (2011).
КАС
пабмед
СтатьяGoogle ученый
Fersht, A. R. Structure and Mechanism in Protein Science (Freeman, 1999).
Хенцлер-Вилдман К. и Керн Д. Динамические свойства белков.
Природа 450 , 964–972 (2007).КАС
пабмед
СтатьяGoogle ученый
Yao, Y., Belcher, J., Berger, A.J., Mayer, M.L. & Lau, A.Y. Конформационный анализ лиганд-связывающих доменов GluN1, GluN2 и GluN3 рецепторов NMDA выявил специфические для подтипа характеристики. Структура 21 , 1788–1799 (2013).
КАС
пабмед
СтатьяGoogle ученый
Ландес, С. Ф., Рамбхадран, А., Тейлор, Дж. Н., Салатан, Ф. и Джаяраман, В. Структурный ландшафт изолированных агонист-связывающих доменов из отдельных рецепторов AMPA. Нац. хим. биол. 7 , 168–173 (2011).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Olofsson, L. et al. Тонкая настройка субмиллисекундной конформационной динамики контролирует эффективность агонистов метаботропных рецепторов глутамата.
Нац. коммун. 5 , 5206 (2014).КАС
пабмед
СтатьяGoogle ученый
Долино, Д. М., Резаи Адариани, С., Шейх, С. А., Джаяраман, В. и Санабрия, Х. Конформационный отбор и субмиллисекундная динамика лиганд-связывающего домена рецептора N-метил-d-аспартата. Дж. Биол. хим. 291 , 16175–16185 (2016).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Ahmed, A.H., Loh, A.P., Jane, D.E. & Oswald, R.E. Динамика глутамат-связывающего домена S1S2 GluR2, измеренная с помощью 19F ЯМР-спектроскопии. Дж. Биол. хим. 282 , 12773–12784 (2007).
КАС
пабмед
СтатьяGoogle ученый
Джин, Р., Банке, Т. Г., Майер, М. Л., Трайнелис, С. Ф. и Гуо, Э. Структурная основа частичного агонистического действия на ионотропные глутаматные рецепторы.
Нац. Неврологи. 6 , 803–810 (2003).КАС
пабмед
СтатьяGoogle ученый
Рамасвами, С. и др. Роль конформационной динамики в частичном агонизме рецептора альфа-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты (АМРА). Дж. Биол. хим. 287 , 43557–43564 (2012).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Ахмед, А. Х., Ван, С., Чуанг, Х. Х. и Освальд, Р. Э. Механизм активации рецептора AMPA частичными агонистами: дисульфидное улавливание конформаций с закрытыми лепестками. Дж. Биол. хим. 286 , 35257–35266 (2011).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Paas, Y. Макро- и микроархитектуры лиганд-связывающего домена глутаматных рецепторов.
Тренды Неврологии. 21 , 117–125 (1998).КАС
пабмед
СтатьяGoogle ученый
Paas, Y., Eisenstein, M., Medevielle, F., Teichberg, V.I. & Devillers-Thiery, A. Идентификация подмножеств аминокислот, отвечающих за специфичность связывания лиганда рецептором глутамата. Нейрон 17 , 979–990 (1996).
КАС
пабмед
СтатьяGoogle ученый
Durham, R. J. et al. Конформационное распространение и динамика аллостерии NMDA-рецепторов. Проц. Натл акад. науч. США 117 , 3839–3847 (2020).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Шулер, Б. и Хофманн, Х. Спектроскопия одиночных молекул динамики сворачивания белков — расширение масштабов и временных масштабов. Курс.
мнение Структура биол. 23 , 36–47 (2013).КАС
пабмед
СтатьяGoogle ученый
Бенвенист М., Клементс Дж., Выклицкий Л. мл. и Майер М. Л. Кинетический анализ модуляции рецепторов N-метил-D-аспарагиновой кислоты глицином в культивируемых нейронах гиппокампа мыши. J. Physiol. 428 , 333–357 (1990).
КАС
пабмед
ПабМед Центральный
СтатьяGoogle ученый
Рис, Дж., Шварце, С., Джонсон, С.М. и Нойвейлер, Х. Микросекундная укладка и доменные движения структурного переключателя белка шелка паука. Дж. Ам. хим. соц. 136 , 17136–17144 (2014).
КАС
пабмед
СтатьяGoogle ученый
Кричевский О. и Боннет Г. Корреляционная спектроскопия флуоресценции: метод и его применение.
Рем. прог. физ. (Великобритания) 65 , 251–297 (2002).КАС
СтатьяGoogle ученый
Muller, C.B. et al. Точное измерение диффузии с помощью многоцветной двухфокусной флуоресцентной корреляционной спектроскопии. Epl—Europhys. лат. 83 , 46001 (2008 г.).
Burchard, W., Schmidt, M. & Stockmayer, WH Информация о полидисперсности и разветвлениях от комбинированного квазиупругого и интегрального рассеяния. Макромолекулы 13 , 1265–1272 (1980).
КАС
СтатьяGoogle ученый
Hulme, E.C. & Trevethick, M.A. Анализы связывания лигандов в равновесии: проверка и интерпретация. Брит. Дж. Фарм. 161 , 1219–1237 (2010).
КАС
СтатьяGoogle ученый
Pohlsgaard, J., Frydenvang, K., Madsen, U. & Kastrup, JS. Уроки из более чем 80 структур лиганд-связывающего домена GluA2 в комплексе с агонистами, антагонистами и аллостерическими модуляторами. Нейрофармакология 60 , 135–150 (2011).
КАС
пабмед
Статья
Google ученый
Соболевский А.И. Строение и гейтирование тетрамерных рецепторов глутамата. J. Physiol. 593 , 29–38 (2015).
КАС
пабмед
Статья
Google ученый
Meyerson, J.R. et al. Структурные основы десенсибилизации глутаматных рецепторов каинатного подтипа. Природа 537 , 567–571 (2016).
КАС
пабмед
ПабМед Центральный
Статья
Google ученый
Sun, Y. et al. Механизм десенсибилизации глутаматных рецепторов. Природа 417 , 245–253 (2002).
КАС
пабмед
Статья
Google ученый
Ссылки на скачивание
Благодарности
Авторы благодарят Hiro Furukawa за предоставление плазмид для гетерологичной экспрессии LBD GluN1 и GluN2A. Эта публикация была поддержана Фондом публикаций открытого доступа Вюрцбургского университета.
Финансирование
Финансирование открытого доступа организовано и разрешено Projekt DEAL.
Информация об авторе
Авторы и организации
Кафедра биотехнологии и биофизики, Университет Юлиуса-Максимилиана, Вюрцбург, Вюрцбург, Германия
Suhaila Rajab, Leah Bismin, Simone Schwarze & Hannes Neuweiler
Отдел нейробиологии, Лаборатория медицинских исследований молекулярной биологии, Cambridge, UK
Alexandra Pinggera & Ingo H. Greger
2232 Authorors
7777777777777777777777777777777777777797779779779.j.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.J.JALORS.
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar
GregerПосмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar
Contributions
S.R., S.S., I.H.G. и HN разработали эксперименты. С.Р., С.С., Л.Б. и А.П. проводили эксперименты. С.Р., С.С. и Л.Б. проанализированные данные. Статью написали С. Р., А. П., И. Х. Г. и Х. Н..
Автор, ответственный за переписку
Связь с
Ханнес Нойвейлер.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Информация о рецензировании Биология коммуникаций благодарит Роберта Кваста и других анонимных рецензентов за их вклад в рецензирование этой работы. Основные редакторы обработки: Кришнананда Чаттопадхьяй и Анам Ахтар. Доступны отчеты рецензентов.
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Supplementary information
Transparent Peer Review File
Supplementary Information
Description of Supplementary Files
Supplementary Data 1
Reporting summary
Rights and permissions
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала. и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или превышает разрешенное использование, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Механизм диссоциации процессивных целлюлаз
1. Payne C.M., et al., Fungal Cellulases. хим. преп.
115, 1308–1448 (2015). [PubMed] [Google Scholar]
2. Крэгг С. М. и др., Механизмы деградации лигноцеллюлозы на древе жизни. Курс. мнение хим. биол.
29, 108–119 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. Клемм Д., Хойблейн Б., Финк Х. П., Бон А., Целлюлоза: увлекательный биополимер и устойчивое сырье. Ангью. хим. Междунар. Эд. англ.
44, 3358–3393 (2005). [PubMed] [Google Scholar]
4. Вольфенден Р. Степени сложности водопотребляющих реакций в отсутствие ферментов. хим. преп.
106, 3379–3396 (2006). [PubMed] [Google Scholar]
5. Chundawat S.P.S., Beckham G.T., Himmel M.E., Dale B.E., Разложение лигноцеллюлозной биомассы на топливо и химикаты. Анну. Преподобный Хим. биомол. англ.
2, 121–145 (2011). [PubMed] [Академия Google]
6. Химмель М. Е. и др., Устойчивость биомассы: Инженерные установки и ферменты для производства биотоплива. Наука
315, 804–807 (2007). [PubMed] [Google Scholar]
7. Wolfenden R., Lu X., Young G., Спонтанный гидролиз гликозидов. Варенье. хим. соц.
120, 6814–6815 (1998). [Google Scholar]
8. Ким С., Дейл Б. Е. Распределенная система биопереработки целлюлозы на Среднем Западе США на основе кукурузной соломы. Биотопливо Биопрод. Биорефин.
10, 819–832 (2016). [Академия Google]
9. Wilson D.B. Целлюлазы и биотопливо. Курс. мнение Биотехнолог.
20, 295–299 (2009). [PubMed] [Google Scholar]
10. Дейл Б., Время переосмыслить целлюлозное биотопливо?
Биотопливо Биопрод. Биорефин.
12, 5–7 (2018). [Google Scholar]
11. Вальдивия М., Галан Дж. Л., Лаффарга Дж., Рамос Дж. Л., Биотопливо 2020: Биоперерабатывающие заводы на основе лигноцеллюлозных материалов. микроб. Биотехнолог.
9, 585–594 (2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Перальта-Яхья П. П., Чжан Ф., дель Кардайр С. Б., Кислинг Дж. Д., Микробиологическая инженерия для производства современного биотоплива. Природа
488, 320–328 (2012). [PubMed] [Академия Google]
13. Серрано-Руис Дж. К., Думесик Дж. А., Каталитические пути преобразования биомассы в жидкие углеводородные транспортные топлива. Энергетическая среда. науч.
4, 83–99 (2011). [Google Scholar]
14. Momeni M.H., et al., Структурная, биохимическая и компьютерная характеристика гликозидгидролазы семейства 7 целлобиогидролазы грибка-убийцы деревьев Heterobasidion lineare . Дж. Биол. хим.
288, 5861–5872 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Ялак Й., Курашин М., Теугьяс Х., Вялямяэ П., Эндо-экзосинергизм в гидролизе целлюлозы. Дж. Биол. хим.
287, 28802–28815 (2012). [PMC free article] [PubMed] [Google Scholar]
16. Курасин М., Вялямяэ П. Процессивность целлобиогидролаз ограничена субстратом. Дж. Биол. хим.
286, 169–177 (2011). [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Cruys-Bagger N., Tatsumi H., Ren G.R., Borch K., West P., Переходная кинетика и этапы ограничения скорости для процессивной целлобиогидролазы Cel7A: Влияние структуры субстрата и углеводсвязывающего домена. Биохимия
52, 8938–8948 (2013). [PubMed] [Google Scholar]
18. Накамура А. и др. Компромисс между процессивностью и гидролитической скоростью целлобиогидролаз на поверхности кристаллической целлюлозы. Варенье. хим. соц.
136, 4584–4592 (2014). [PubMed] [Google Scholar]
19. Kipper K., Väljamäe P., Johansson G. Процессуальное действие целлобиогидролазы Cel7A из Trichoderma reesei проявляется в виде кинетики «взрыва» на флуоресцентных полимерных модельных субстратах. Биохим. Дж.
385, 527–535 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Ståhlberg J., Johansson G., Pettersson G., Trichoderma reesei не имеет настоящей экзоцеллюлазы: все интактные и укороченные целлюлазы продуцируют новые редуцирующие концевые группы на целлюлозе. Биохим. Биофиз. Акта
1157, 107–113 (1993). [PubMed] [Google Scholar]
21. Schiano-di-Cola C., et al., Систематические делеции в целлобиогидролазе (CBH) Cel7A гриба Trichoderma reesei обнаруживают гибкие петли, критически важные для активности CBH. Дж. Биол. хим.
294, 1807–1815 (2019 г.)). [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Бадино С. Ф. и др., Экзо-экзо синергия между Cel6A и Cel7A из Hypocrea jecorina : Роль модуля связывания углеводов и эндолитический характер ферменты. Биотехнолог. биоинж.
114, 1639–1647 (2017). [PubMed] [Google Scholar]
23. Knott B.C., Crowley M.F., Himmel M.E., Ståhlberg J., Beckham G.T., Взаимодействия углевод-белок, которые управляют процессуальной транслокацией полисахаридов в ферментах, выявленные в результате компьютерного исследования процессивности целлобиогидролазы. Варенье. хим. соц.
136, 8810–8819(2014). [PubMed] [Google Scholar]
24. Knott B.C., et al., Механизм гидролиза целлюлозы с помощью двухстадийной удерживающей целлобиогидролазы, выясненный путем изучения структурных и переходных путей. Варенье. хим. соц.
136, 321–329 (2014). [PubMed] [Google Scholar]
25. Кошланд Д. Э. мл. Стереохимия и механизм ферментативных реакций. биол. Преподобный Кэмб. Филос. соц.
28, 416–436 (1953). [Google Scholar]
26. Bu L., et al., Изучение вытеснения углеводного продукта из процессивной целлюлазы с помощью нескольких методов абсолютной связывающей свободной энергии. Дж. Биол. хим.
286, 18161–18169(2011). [Статья бесплатно PMC] [PubMed] [Google Scholar]
27. Kari J., et al., Кинетика вариантов целлобиогидролазы (Cel7A) с пониженным сродством к субстрату. Дж. Биол. хим.
289, 32459–32468 (2014). [Статья бесплатно PMC] [PubMed] [Google Scholar]
28. Cruys-Bagger N., et al., Предустановившаяся кинетика гидролиза нерастворимой целлюлозы целлобиогидролазой Cel7A. Дж. Биол. хим.
287, 18451–18458 (2012). [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Igarashi K., et al., Пробки снижают гидролитическую эффективность целлюлазы на поверхности целлюлозы. Наука
333, 1279–1282 (2011). [PubMed] [Google Scholar]
30. Vermaas J. V., et al., Механизм ингибирования лигнином ферментативной деконструкции биомассы. Биотехнолог. Биотопливо
8, 217 (2015). [Статья бесплатно PMC] [PubMed] [Google Scholar]
31. Shibafuji Y., et al., Одномолекулярный анализ элементарных стадий реакции Trichoderma reesei целлобиогидролазы I (Cel7A), гидролизующей кристаллическую целлюлозу I α и III I . Дж. Биол. хим.
289, 14056–14065 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Соренсен Т. Х., Крюйс-Баггер Н., Борх К., Вест П. Диаграмма свободной энергии для гетерогенного ферментативного гидролиза гликозидных связей в целлюлозе. Дж. Биол. хим.
290, 22203–22211 (2015). [Статья бесплатно PMC] [PubMed] [Google Scholar]
33. Jalak J., Väljamäe P., Механизм начального быстрого замедления скорости гидролиза целлюлозы, катализируемого целлобиогидролазой. Биотехнолог. биоинж.
106, 871–883 (2010). [PubMed] [Google Scholar]
34. Cruys-Bagger N., Elmerdahl J., Praestgaard E., Borch K., West P., Теория стационарного состояния процессивных целлюлаз. ФЕБС Дж.
280, 3952–3961 (2013). [PubMed] [Google Scholar]
35. Шультен К., Шультен З., Сабо А., Динамика реакций при пересечении диффузионного барьера. Дж. Хим. физ.
74, 4426–4432 (1981). [Google Scholar]
36. Ståhlberg J., Knott B.C., запись PDB 6RWF. Банк данных о белках. https://www.rcsb.org/structure/6RWF. Депонировано 4 июня 2019 г.
37. Курашин М., Кууск С., Кууск П., Сёрли М., Вялямяэ П., Медленные скорости диссоциации и сильное связывание продукта необходимы для процессивности и эффективного разложения неподатливого хитина семейством 18. хитиназы. Дж. Биол. хим.
290, 29074–29085 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Kont R., Kari J., Borch K., Westh P., Väljamäe P., Междоменный синергизм необходим для эффективной подачи целлюлозной цепи в активную сайт целлобиогидролазы Cel7A. Дж. Биол. хим.
291, 26013–26023 (2016). [Статья бесплатно PMC] [PubMed] [Google Scholar]
39. Sørensen T.H., et al., Варианты петли термофила Rasamsonia emersonii Cel7A с улучшенной активностью против целлюлозы. Биотехнолог. биоинж.
114, 53–62 (2017). [PubMed] [Академия Google]
40. Vermaas J.V., Crowley M.F., Beckham G.T., Payne C.M. Влияние окисления литического полисахарида монооксигеназой на структуру целлюлозы и связывание олигомеров окисленной целлюлозы с целлюлазами. Дж. Физ. хим. Б
119, 6129–6143 (2015). [PubMed] [Google Scholar]
41. Silveira R.L., Skaf M.S. Молекулярно-динамическое моделирование мутантов целлобиогидролазы семейства 7, направленное на снижение ингибирования продукта. Дж. Физ. хим. Б
119, 9295–9303 (2015). [PubMed] [Академия Google]
42. Сильвейра Р.Л., Скаф М. С., Согласованные движения и крупномасштабные структурные колебания целлобиогидролазы Trichoderma reesei Cel7A. физ. хим. хим. физ.
20, 7498–7507 (2018). [PubMed] [Google Scholar]
43. Brady S.K., Sreelatha S., Feng Y., Chundawat S.P.S., Lang M.J., Целлобиогидролаза 1 из Trichoderma reesei разлагает целлюлозу за один шаг. Нац. коммун.
6, 10149 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Beckham G.T., et al., К теории молекулярного уровня процессивности углеводов в гликозидгидролазах. Курс. мнение Биотехнолог.
27, 96–106 (2014). [PubMed] [Google Scholar]
45. Лютербахер Дж. С., Уокер Л. П., Моран-Мирабаль Дж. М., Наблюдение и моделирование деградации BMCC с помощью коммерческих коктейлей целлюлаз с флуоресцентно меченым Trichoderma reseii Cel7A с помощью конфокальной микроскопии. Биотехнолог. биоинж.
110, 108–117 (2013). [PubMed] [Google Scholar]
46. Kari J., et al., Аномерная селективность и профиль продукта процессивной целлюлазы. Биохимия
56, 167–178 (2017). [PubMed] [Академия Google]
47. Westh P., et al., Термоактивация целлобиогидролазы. Биотехнолог. биоинж.
115, 831–838 (2018). [PubMed] [Google Scholar]
48. Igarashi K., et al., Высокоскоростная атомно-силовая микроскопия визуализирует процессное движение Trichoderma reesei целлобиогидролазы I на кристаллической целлюлозе. Дж. Биол. хим.
284, 36186–36190 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Нидецкий Б., Захария В., Геркен Г., Хайн М., Штайнер В., Гидролиз целлоолигосахаридов с помощью1363 Trichoderma reesei целлобиогидролазы: экспериментальные данные и кинетическое моделирование. Ферментный микроб. Технол.
16, 43–52 (1994). [Google Scholar]
50. Harjunpää V., et al., Гидролиз целлоолигосахаридов целлобиогидролазой II из Trichoderma reesei . Константы ассоциации и скорости, полученные на основе анализа кривых прогресса. Евро. Дж. Биохим.
240, 584–591 (1996). [PubMed] [Google Scholar]
51. Рабинович М. Л., Мельник М. С., Хернер М. Л., Возный Ю. В., Васильченко Л. Г. Преимущественное непродуктивное связывание субстрата грибковой целлобиогидролазой I и значение для улучшения активности. Биотехнолог. Дж.
14, e1700712 (2019 г.). [PubMed] [Google Scholar]
52. Кари Дж. и др. Принцип Сабатье для межфазного (гетерогенного) ферментативного катализа. Катал.
8, 11966–11972 (2018). [Google Scholar]
53. von Ossowski I., et al., Разработка экзо-петли целлобиогидролазы Trichoderma reesei , Cel7A. Сравнение с Phanerochaete chrysosporium Cel7D. Дж. Мол. биол.
333, 817–829 (2003). [PubMed] [Google Scholar]
54. Чжан С., Ирвин Д. К., Уилсон Д. Б., Сайт-направленная мутация некаталитических остатков Thermobifida fusca экзоцеллюлаза Cel6B. Евро. Дж. Биохим.
267, 3101–3115 (2000). [PubMed] [Google Scholar]
55. Бекхэм Г. Т. и др. Происхождение непокорности биомассы на молекулярном уровне: свободная энергия декристаллизации для четырех распространенных полиморфов целлюлозы. Дж. Физ. хим. Б
115, 4118–4127 (2011). [PubMed] [Google Scholar]
56. Payne C.M., et al., Процессивность гликозидгидролазы напрямую связана со свободной энергией связывания олигосахаридов. Варенье. хим. соц.
135, 18831–18839 гг.(2013). [PubMed] [Google Scholar]
57. Брандт А., Грасвик Дж., Халлетт Дж. П., Велтон Т., Деконструкция лигноцеллюлозной биомассы с помощью ионных жидкостей. Зеленый хим.
15, 550–583 (2013). [Google Scholar]
58. Хорн С. Дж. и др. Затраты и преимущества процессивности при ферментативной деградации неподатливых полисахаридов. проц. Натл. акад. науч. США.
103, 18089–18094 (2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Брукс Б. Р. и др., CHARMM: Программа биомолекулярного моделирования. Дж. Комп. хим.
30, 1545–1614 (2009 г.)). [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Phillips J.C., et al., Масштабируемая молекулярная динамика с NAMD. Дж. Комп. хим.
26, 1781–1802 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. День А. и др., «Вариант целлюлазы Hypocrea jecorina CBh2». Патент США 8 637 294 B2 (2014 г.).
62. Momeni M.H., et al., Экспрессия, кристаллическая структура и целлюлазная активность термостабильной целлобиогидролазы Cel7A из гриба Humicola grisea var. термоидея. Акта Кристаллогр. Д биол. Кристаллогр.
70, 2356–2366 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Ståhlberg J., et al., Исследования активности и кристаллической структуры каталитически дефицитных мутантов целлобиогидролазы I из Trichoderma reesei . Дж. Мол. биол.
264, 337–349 (1996). [PubMed] [Google Scholar]
64. Goedegebuur F., et al., Улучшение термостабильности целлобиогидролазы Cel7A из Hypocrea jecorina путем направленной эволюции. Дж. Биол. хим.
292, 17418–17430 (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Momeni M.H., Ubhayasekera W., Sandgren M., Ståhlberg J., Hansson H., Структурное понимание ингибирования целлобиогидролазы Cel7A ксилоолигосахаридами. ФЕБС Дж.
282, 2167–2177 (2015). [PubMed] [Академия Google]
66. Таунс Дж. и др., XSEDE: Ускорение научных открытий. вычисл. науч. англ.
16, 62–74 (2014). [Google Scholar]
67. Аморе А. и др. Различная роль N — и O -гликанов в активности и стабильности целлюлазы. проц. Натл. акад. науч. США.
114, 13667–13672 (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Грейферы-раскладушки — BLUG
Грейферы-раскладушки — BLUG
Откройте для себя полный ассортимент грейферных грейферов, которые мы предлагаем в зависимости от механизма привода и плотности, с которыми вам необходимо работать: сыпучие материалы и продукты, удобрения, шлам, навоз, песок, щебень, камни, шелуха, цемент, горячий клинкер…
Грейфер грейферный электрогидравлический
Грейфер грейферный электрогидравлический состоит из: электродвигателя, насоса и электрогидравлических клапанов, обеспечивающих давление в системе и корпусе грейфера, являющихся всеми элементами, собранными в корпусе машины.
С2-1,1
Свайный и сыпучий материал плотностью до 1,1 т/м³.
С2-1,8
Свайный и сыпучий материал плотностью до 1,8 т/м³.
С2-2,8
Свайный и сыпучий материал плотностью до 2,8 т/м³.
С2Н-1,1
Свайный и сыпучий материал плотностью до 1,1 т/м³.
С2Н-1,8
Свайный и сыпучий материал плотностью до 1,8 т/м³.
С2Н-2,8
Свайный и сыпучий материал плотностью до 2,8 т/м³.
С4-1,1
Свайный и сыпучий материал плотностью до 1,1 т/м³.
С4-1,8
Цемент, горячий клинкер (300°С) и свайный и сыпучий материал плотностью до 1,8 т/м³.
С4-2,8
Свайный и сыпучий материал плотностью до 2,8 т/м³.
ЦВ2-1,1
Свайный и сыпучий материал плотностью до 1,1 т/м³.
ЦВ2-1,8
Свайный и сыпучий материал плотностью до 1,8 т/м³.
ЦВ2-2,8
Свайный и сыпучий материал плотностью до 2,8 т/м³.
Механические грейферные грейферы
Грейферные грейферы, приводимые в действие и подвешенные на тросах. Существуют три весы: одноканатные, двухканатные и четырехканатные грейферы.
СМ4-0,8
Удобрения, наваленные и сыпучие материалы плотностью до 0,8 т/м³.
СМ4-1,1
Удобрения, злаки, уголь, свалки и сыпучие материалы плотностью до 1,1 т/м³.
СМ1-1,8
Удобрения, крупы, уголь, цемент, клинкер, свайные и сыпучие материалы плотностью до 1,8 т/м³.
СМ4-2,8
Полезные ископаемые, насыпные и сыпучие материалы плотностью до 2,8 т/м³.
СМ4-3,2
Полезные ископаемые, наваленные и сыпучие материалы плотностью до 3,2 т/м³.
CM4G-0,8
Удобрения, наваленные и сыпучие материалы плотностью до 0,8 т/м³.
CM4G-1,1
Удобрения, злаки, уголь, свалки и сыпучие материалы плотностью до 1,1 т/м³.
СМ1Р-1,8
Цемент, клинкер, бокситы, полевошпатовые, свайные и сыпучие материалы плотностью до 1,8 т/м³.
СМ2-1,8
Цемент, клинкер, свайный и сыпучий материал плотностью до 1,8 т/м³.
СМ4-1,8
Цемент, клинкер, свайный и сыпучий материал плотностью до 1,8 т/м³.
Грейферы гидравлические
Грейферы неавтономные. Приводится в движение гидравлически группой посторонних грейферов. Масло под давлением должно поступать извне.
ЦНАХ-2,8
Камни, гравий, песок, ил, подводные отходы в дренажных выработках, свайный и сыпучий материал плотностью до 2,8 т/м³.
Экологически чистые захваты
Пример открытого клапана: Падение материала при транспортировке грейфером
Пример закрытого клапана: экологически чистый захват
- Экологически безопасные модели исключают любой риск падения материала во время транспортировки.
- Двойное эластичное закрытие между кромками клапанов для материалов в порошке .
Проектирование и моделирование
- Наше конструкторское бюро разрабатывает каждую конструкцию в соответствии с F.E.M. 1001 Раздел I:1998, UNE-58132 и UNE-EN 13155 с использованием новейших инструментов 3D и CAD/CAM.
- Каждая новая конструкция проверяется и оптимизируется с использованием метода анализа конечных элементов для анализа эффектов усталости и гарантии 2 000 000 циклов на срок службы компонентов и машин.
Функционирование электрогидравлической системы
| Реверс двигателя | Клапан с электроприводом |
|---|---|
| Количество проводов: 4 | Количество проводов: 7 |
| Тип насоса: Поршневой насос (фиксированный расход) | Тип насоса: Шестеренчатый насос (постоянный поток) Поршневой насос (постоянный поток) Поршневой насос (переменный поток) |
Гидравлические агрегаты с переменным расходом
- Гидравлические агрегаты с электрическим клапаном, использующие 9Поршневые насосы с переменным расходом 1678 гарантируют увеличение срока службы гидравлических компонентов более чем на 300% по сравнению с насосами с постоянным расходом.
- Эта система предотвращает перегрев масла (в 4 раза меньше, чем в насосах с постоянным расходом) благодаря управлению с определением нагрузки. Таким образом, плита насоса постоянно адаптирует свой наклон таким образом, чтобы оптимизировать усилие проникновения и минимизировать потребляемую мощность (на 40% меньше, чем у насосов с фиксированным потоком) , а также повысить общую эффективность грейфера.
Материалы
- Конструкция изготовлена из катаной стали S355 J2G3 (упругий предел текучести 510-610 Н/мм2).
- Возможна установка сменных марганцевых износостойких зубьев (до 500 HB) в грейферные клапаны таким образом, чтобы улучшить проникновение машины в материал.
- Для захватов, контактирующих с абразивными материалами мы обычно рекомендуем изготавливать пластины клапанов из износостойкая сталь (твердость до 475 HBW).
Сертификация Atex
Возможность сертификации любой из наших моделей в зоне ATEX 1/21 или 2/22 для взрывоопасных сред.
Свяжитесь с нами
Файлы cookie на этом веб-сайте используются для персонализации контента и рекламы, предоставления функций социальных сетей и анализа трафика.
Настроить файлы cookie
Принять файлы cookie
Лучших грейферных термопрессов [25 машин + обзоры]
Сортировать по:
РекомендованныеЛучшие продажиПо алфавиту: A-ZПо алфавиту: Z-AЦена: от низкой до высокойЦена: от высокой до низкойДата: от новой до старойДата: от старой к новой
В термопрессах с грейферным корпусом используется механизм с верхней и нижней пластинами, которые напоминают раковину моллюска или челюсти. Они обычно дешевле, прочнее и занимают меньше места. Они бывают всех размеров от меньше чем 15×15 до 20×25 дюймов. Когда вы нажимаете на одежду, верхняя плита опускается с помощью шарнирного механизма. Альтернативой раскладной конструкции является откидной термопресс. Они требуют больше места, но более безопасны и просты в эксплуатации.
+ Быстрый магазин
Rincons 15 x 15 дюймов Машина для термопрессования раскладушки AP4-15LCD
Распроданный
$399,00
Rincons 15 x 15 дюймов AP4-15LCD, грейферная машина для термопрессования
Распроданный
$399,00
Этот коммерческий термопресс 15×15 имеет большой элемент 15″ x 15″ (фактический размер 38см x 38см), который позволяет вам создавать нестандартные футболки, номерные знаки и многие другие уникальные. ..
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Ricoma iKonix Flat 15″ x 15″ грейферная машина HP-3838H с термопрессом
$389,00
$429.00
Ricoma iKonix Flat 15″ x 15″ грейферный термопресс HP-3838H
$389,00
429,00 долларов США
Тепловой пресс iKonix 15 x 15 дюймов идеально подходит для переноса больших объемов футболок. Этот раскладной термопресс лучше всего подходит для начинающих, которым нужна надежная термопресс-машина. …
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Rincons 16 x 20 дюймов Машина для переноса термопресса раскладушки AP-V20 портретной ориентации
Продано
Rincons 16 дюймов x 20 дюймов AP-V20, портретная термопресс-трансферная машина
Распроданный
Поверхность нагрева с антипригарным покрытием. Возможность загрузки под высоким давлением. Цифровой дисплей таймер и датчик температуры. Конструкция стальной рамы, очень толстые соединения и стержни компонентов конструкции. Легкое и широкое открывание. 16…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Rincons 16 x 20 дюймов Машина для переноса термопресса раскладушки AP4-20
Продано
Rincons 16 x 20 дюймов грейферный термопресс AP4-20
Распроданный
Этот коммерческий термопресс имеет большой элемент 16″ x 20″ (фактический размер 15-3/4″ x 19″). -3/4″), что позволяет создавать собственные футболки, номерные знаки и многие другие уникальные и…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Ricoma iKonix Flat 16″ x 24″ грейферная машина HP-6040H с термопрессом
Распроданный
$479,00
Ricoma iKonix Flat 16″ x 24″ грейферный термопресс HP-6040H
Распроданный
479,00 долларов США
Тепловой пресс iKonix 16 x 24 дюйма идеально подходит для переноса больших объемов футболок. Раскладные термопрессы лучше всего подходят для дизайнеров одежды, которым нужна надежная машина. Функции:…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Rincons 16 x 24 дюйма Ландшафтная раскладушка AP2-24 Машина для термопрессования
Распроданный
$729,00
Rincons 16″ x 24″ Пейзажная раскладушка AP2-24 Машина для термопрессования
Распроданный
729,00 долларов США
Это горизонтальный термопресс размером 16×24 дюйма. Каркас из прочной стали весом 130 фунтов. Рама рукоятки длиннее, чтобы обеспечить больший рычаг и крутящий момент при минимальных усилиях. Этот…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 15 x 15 дюймов Clamshell HT-400E Машина для термопрессования
$901,25
HIX 15 x 15 дюймов грейферный термопресс HT-400E
$901,25
Термопресс-трансфер Clamshell HT-400E — один из самых экономичных, удобных и простых в эксплуатации термопрессов HIX. Он идеально подходит для опытных мастеров и начинающих графических бизнесменов, а также…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Geo Knight 14 x 16 дюймов Clamshell DK16 Машина для термопрессования
Распроданный
1200,00 долларов США
Geo Knight 14 x 16 дюймов грейферный термопресс DK16
Распроданный
1200,00 долларов США
DK16 размером 14 x 16 дюймов открывается для полного доступа к нижнему нижнему столу. Задняя часть нагревательной плиты поднимается над нижним столом и удаляется от него, обеспечивая лучший доступ…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Geo Knight 14 дюймов x 16 дюймов Автоматическая машина для переноса термопресса DK16A Clamshell
Продано
Geo Knight 14 x 16 дюймов Автоматический грейферный термопресс DK16A
Распроданный
DK16A — это AUTO-RELEASE «Pop-Up» версия нашей ручной раскладушки среднего размера. Благодаря этой удивительной высокотехнологичной функции ваша раскладушка Digital Knight работает как полуавтоматический пресс с пневматическим приводом, без…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Geo Knight 16 x 20 дюймов Clamshell DK20 Машина для термопрессования
Продано
Geo Knight 16 x 20 дюймов грейферный термопресс DK20
Распроданный
Наша самая большая раскладушка DK20 16×20 открывается для полного доступа к нижнему нижнему столу. Важно отметить, что задняя часть нагревательной плиты поднимается выше и…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 15 x 15 дюймов Машина для термопрессования грейферного типа HT-400
1475,00 долларов США
HIX 15 x 15 дюймов, грейферный термопресс HT-400
1475,00 долларов США
Графическая индустрия называет эту машину «Рабочей лошадкой» по уважительной причине — грейферный печатный станок HIX HT-400 прочный, долговечный и создан для работы. Необходимая вещь для любого серьезного стартапа…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Geo Knight 16 x 20 дюймов Автоматическая выдвижная раскладушка DK20A Машина для термопрессования
Продано
Geo Knight 16 x 20 дюймов Автоматическая выдвижная грейферная машина DK20A Термопресс для трансфера
Распроданный
DK20A — это AUTO-RELEASE «Pop-Up» версия нашей самой большой ручной раскладушки. Благодаря этой удивительной высокотехнологичной функции ваша раскладушка Digital Knight работает как полуавтоматический пресс с пневматическим приводом, без…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 16 x 20 дюймов HT-600 Машина для переноса тепла с грейферным корпусом
$1675,00
HIX 16 x 20 дюймов, грейферный термопресс HT-600
1675,00 долларов США
Пресс HIX HT-600 Clamshell Press гарантирует, что ваши большие оттиски с термопереносом будут идеальными от края до края. Благодаря большей поверхности плиты размером 16 x 20 дюймов вы сможете…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 15 x 15 дюймов Термопресс-трансферная машина S-450 с автоматически открывающейся грейферной оболочкой
$1795,00
HIX 15 x 15 дюймов Термопресс для переноса грейферного корпуса S-450 с автоматическим открытием
1795,00 долларов США
Закройте крышку и уходите. Найди другую работу или возьми газировку. С теплообменной машиной HIX S-450 с автоматическим выпуском у вас будет беспроблемная работа. Когда…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Insta 15 x 20 дюймов, модель 158, плоский грейферный термопресс с автоматическим выпуском
1830,00 долларов США
Плоский грейферный термопресс Insta 15 x 20 дюймов, модель 158, с автоматическим выпуском
1830,00 долларов США
Термопресс Insta Model 158 Digital Auto Release предлагает одно из основных преимуществ автоматической машины — функцию автоматического открытия — в легком, недорогом, эргономичном дизайне. ..
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 16 x 20 дюймов S-650 Автоматически открывающаяся грейферная машина для термопереноса
1 995,00 долларов США
HIX 16 x 20 дюймов S-650 Термопресс для автоматического открывания грейферного пресса
1995,00 долларов США
Получите покрытие, необходимое для ваших проектов, с автоматически открывающимся грейферным термопрессом HIX S-650. Этот выдающийся пресс открывается автоматически без использования воздушного компрессора. Это идеальный пресс…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
Geo Knight 14 x 16 дюймов Многоцелевой термопресс DC16AP
Продано
Многоцелевой термопресс Geo Knight 14 x 16 дюймов DC16AP
Распроданный
Geo Knight DC16AP — это пневматический автоматический многоцелевой откидной термопресс размером 14 x 16 дюймов с дополнительными сменными нагревательными плитами и столами. Плавное автоматическое нажатие и активация кнопки обеспечивают…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 15 x 15 дюймов Пневматическая грейферная машина N-680 с термопрессом
2835,00 долларов США
HIX 15 x 15 дюймов Пневматический грейферный термопресс N-680
2835,00 долларов США
Пневматические автоматические термопрессы HIX N-680 можно описать тремя фразами: простота использования, высокая эффективность и надежность. Вы заметите, что ваши операторы будут меньше уставать и…
Посмотреть полную информацию о продукте »
+ Быстрый магазин
HIX 16 x 20 дюймов Пневматическая грейферная машина N-880 с термопрессом
3 655,00 долларов США
HIX 16 x 20 дюймов Пневматический грейферный термопресс N-880
3655,00 долларов США
Автоматический термопресс HIX N-880 с пневматическим приводом предоставляет ряд возможностей, на которые вы можете положиться. Благодаря эффективности, согласованности и надежной производительности у вас будет…
Посмотреть полную информацию о продукте »
| ||||||||||||
