

Содержание
МАЗ-5336 технические характеристики, двигатель и расход топлива, кабина, размеры и отзывы
Автомобиль «МАЗ-5336» – одна из получивших самое широкое распространение на постсоветском пространстве моделей Минского автомобильного завода. Это универсальное среднетоннажное двухосное шасси, максимальная техническая грузоподъемность которого установлена в 10,9 тонн. Колёсная формула – 4х2. С успехом используется для установки разнообразного специального оборудования и надстроек. Однако основным применением семейства стало использование в составе автопоезда, на «дальнобойных» грузоперевозках.
Об истории модели
Период с 1981 по 1988 годы стал для Минского автозавода временем широкомасштабной реконструкции, когда была полностью обновлена вся линейка выпускаемой продукции. Новые кабины с откидными решётками передней панели и другой приборной доской; модернизированные передние оси, задние мосты, иные рама и тормозная система… С конца 80-х годов начался период серийного производства обновлённых, модернизированных семейств «МАЗ»ов, в числе основных из них – и «МАЗ-5336».
Данную модель можно назвать в линейке предприятия преемником «МАЗ-5335», который стоял на конвейере Минского автомобильного завода с 1977-го года. Это также одно из базовых семейств предприятия, поэтому его изменения прорабатывались особенно тщательно. Новый автомобиль «МАЗ-5336» уже полностью соответствовал современным нормам Международной таможенной системы «T.I.R.», предусматривающей транспортировку грузов на транзитных перевозках, в том числе и в составе автопоезда.
Главными отличиями «МАЗ-5336» от его предшественника стали: набор более мощных двигателей; модернизированная, более мягкая подвеска; увеличенная грузоподъёмность; улучшенные показатели экономичности, усовершенствованные вибро- и шумоизоляция, более просторный грузовой отсек и увеличенная, изменённая в соответствии с велениями времени, кабина. А также наличие уже в базовой комплектации солнцезащитного козырька J. В 1990 году «МАЗ-5335» уже окончательно уступил своё место на конвейере Минского автомобильного .
История создания
Бортовой МАЗ-5336 пришел на смену МАЗ-5335
Шасси и бортовой МАЗ-5336 разрабатывались на базе «старичка» МАЗ-5335, который находился на конвейере с 1977 года. Первые несколько лет оба грузовых автомобиля выпускались параллельно, но, начиная с 1990 года, выпуск МАЗ-5335 был остановлен, и остались только его продвинутые версии — семейство МАЗ-5336.
Нужно отметить, что работа по созданию МАЗ-5336 была масштабная. Взяв за базу 5335-й, инженеры переработали практически все системы грузовика: передние оси, задний мост, раму, рулевое управление, тормозную систему, а также обновили дизайн и предложили новое техническое оснащение.
МАЗ-5336 используется в составе автопоезда
Отличие МАЗ-5336 от МАЗ-5335
От своего предшественника МАЗ-5336 отличается весьма существенно. Если говорить о ключевых различиях, то это:
- линейка более производительных и экономичных моторов;
- увеличенная грузоподъемность;
- более мягкая и пружинистая подвеска;
- просторная кабина;
- улучшенная шумо- и виброизоляция;
- гидропривод для откидывания кабины;
- соответствие международному стандарту T.
 I.R.
I.R.
 I.R.
I.R.МАЗ-5336 получил гидропривод для откидывания кабины
В целом можно отметить, что на момент выпуска МАЗ-5336 грузовик отличался современным и достаточно технологичным оснащением. По эксплуатационным характеристикам он не уступал ни отечественным, ни зарубежным «одноклассникам». При этом в пользу белорусского грузовика были низкая цена и доступность запчастей.
Технические характеристики в цифрах
Габаритные размеры:
- Длина – 8,6 м; ширина – 2,57 м; высота – 3,16 м.
- Колёсная база – 4,9 м;
- Дорожный просвет – 230 мм;
- Минимальный радиус поворота – 9,1 м;
- Площадь платформы – 14.35 кв.метров;
- Внутренние размеры платформы – 6,1 х 2,42 х 2,330 метра;
- Штатный объём стандартной платформы – 34,5 кубических метра.
Параметры массы:
- Масса снаряженного автомобиля – 8,2 тонн;
- Штатная допустимая грузоподъёмность – 9,8 тонн;
- Допустимая полная масса автопоезда – 40 тонн;
- Допустимая полная масса автомобиля – 18 тонн;
- Распределение полной массы: на переднюю ось – 6,5 тонн; на заднюю ось – 11,5 тонн.

Динамические характеристики:
- Максимальная скорость – 100 км/час;
- Время разгона до 60 км/час – 50 секунд;
- Торможение со скорости 60 км/час до нуля – 36,7 метров.
Двигатели
Все модификации грузовиков семейства «МАЗ-5336» оснащались только дизельными двигателями «ЯМЗ» производства Ярославского моторного завода. В линейку агрегатов, устанавливаемых на данную модель грузовика, входят четырёхтактные V-образные восьмицилиндровые, 14,86-литровые, с жидкостным охлаждением и непосредственным впрыском топлива, турбированные (кроме одного) дизельные двигатели.
На «классических» «МАЗ-5336»:
- На «МАЗ-53362» – дизель «ЯМЗ-238Б», номинальная мощность – 300 лошадиных сил.
- На «МАЗ-53361» и «МАЗ-53363» – моторы «ЯМЗ-238ДЕ», номинальная мощность – 330 лошадиных сил.
- На «МАЗ-53366» – НЕтурбированный агрегат «ЯМЗ-238М2», номинальная мощность которого – 240 лошадиных сил.
На «МАЗ-5336» «XXI века»:
- На «МАЗ-5336А3-320» / «МАЗ-5336А5-320» – новые 330-сильные турбированные дизели «ЯМЗ-238ДЕ2», «ЯМЗ-6582. 10Е3» («Евро-3»), с новой же коробкой передач «ЯМЗ-2381(8)».
- На «МАЗ- 5336А3-321» / «МАЗ- 5336А5-321» – новые 250-сильные дизельные моторы «ЯМЗ-6562.10Е3» («Евро-3»), также связке с МКПП «ЯМЗ-2381(8)».
 10Е3» («Евро-3»), с новой же коробкой передач «ЯМЗ-2381(8)».
10Е3» («Евро-3»), с новой же коробкой передач «ЯМЗ-2381(8)».Кроме того, небольшая часть «МАЗ»ов 5336-го семейства (под индексом «МАЗ 533632-321») в начале двухтысячных была выпущена с дизельными двигателями немецкого производства «Дойтц BF 6M1013 FC (Euro-3)», 245 лошадиных сил. Эти моторы работают с коробками передач «ZF 9S75».
Расход топлива
Средний расход топлива для МАЗ-5336 при скорости 60 км/час составляет 21,8 л/100 км. При скорости 80 км/час показатель увеличивается до 29,6 л. В составе автопоезда при скорости 60 км/час на 100 км автомобилю требуется 33,2 л, при скорости 80 км/час – 40,1 л.
Читайте также: Зил 133 гя технические характеристики – ГЯ, Г40, ВЯТ, АЦ-40, Г1, Г, технические характеристики, самосвал, пожарный, тягач, крокодил, тюнинг, лесовоз, автомобиль, цена
Объем топливного бака тягача составляет 255 л.
Трансмиссия грузовика
Трансмиссия «МАЗ-5336» отличается внушительным ресурсом работы и заслуженно считается одним из наиболее надежных элементов конструкции автомобиля данного семейства. Производителем трансмиссии также является Ярославский моторный завод. В составе трансмиссии – двухдисковое сцепление, с гидроусилителем, механическим приводом выключения и периферийными пружинами; 4-х ступенчатая механическая коробка передач «ЯМЗ-2381» с демультипликатором (общее количество передач – восемь). За счёт демультипликатора, повышающего крутящий момент, число передач и увеличивается в 2 раза. Все передачи, за исключением задней, оснащены синхронизаторами.
Карданная передача представлена двумя последовательными валами с промежуточной опорой, главная передача – двухступенчатая и разнесённая. Передаточное число ведущего моста – 5,49.
Подвеска, ходовая часть
Подвеска на автомобилях «МАЗ-5336» – рессорная. Конструкция передней подвески построена на продольных полу-эллиптических рессорах. Передняя подвеска снабжена также амортизаторами и стабилизаторами, обеспечивающими поперечную устойчивость. Задняя подвеска выполнена на двух базовых и двух дополнительных рессорах, и также снабжена стабилизатором поперечной устойчивости.
Передняя подвеска снабжена также амортизаторами и стабилизаторами, обеспечивающими поперечную устойчивость. Задняя подвеска выполнена на двух базовых и двух дополнительных рессорах, и также снабжена стабилизатором поперечной устойчивости.
На грузовиках «МАЗ-5336» установлены бездисковые колеса. Размер шин –«11.00/R20». Рабочая тормозная система имеет барабанные механизмы и двухконтурный пневматический привод. Подобная конструкция сокращает вероятность полного отказа тормозной системы и уменьшает тормозной путь. Стояночный тормоз, естественно, также присутствует в конструкции. Основной прицеп, применяемый для формирования автопоезда с «МАЗ-5336» – модели «МАЗ-837810».
Динамические характеристики
- Максимальная скорость движения автомобиля – 115 км/ч;
- Максимальная скорость движения в составе автопоезда – 100 км/ч;
- Разгон до 60 км/ч – 50 сек;
- Тормозной путь с 60 км/ч – 36,7 м;
- Средний расход топлива на 100 км пути – автомобилем 21,8, автопоездом 33,2;
- Радиус поворота – по внешнему колесу 8,4 м, габаритный 9,1 м.

Электрооборудование
МАЗ 5336 имеет 24-вольтовую систему электропроводки, унифицированную с основными модификациями. В качестве источников электрического тока используются две последовательно соединенные батареи типа 6-СТ182 ЭМ и генераторная установка переменного тока со встроенным выпрямительным блоком и интегрированным регулятором напряжения.
Технические характеристики МАЗ-Купава 5731 (МАЗ-533603)
| Габариты | 8610х2500х4000 мм |
| Кабина | цельнометаллическая трехместная |
| Компоновка | 4х2, задние колеса – ведущие, кабина над двигателем |
| База | 4910 мм |
| Двигатель | дизельный, четырехтактный, V6, ЯМЗ-236 БЕ2 (Евро 2) с турбонаддувом |
| Рабочий объем | 11,2 литров |
| Максимальная мощность | 184,1 кВт 250 л.с. |
| Крутящий момент | 1710 Н*м |
| Коробка передач | восьмиступенчатая механическая с делителем |
| Подвеска спереди и сзади | рессорная на продольных полуэллиптических рессорах |
| Тормоза спереди и сзади | барабанные |
| Максимальная скорость | 105 км/ч |
| Объем бака для топлива | 350 литров |
| Полная масса | 17990 кг |
| Масса перевозимого груза | 10000 кг |
| Внутренние габариты фургона | 6100 х 2486 х 2480 мм |
| Объем кузова | 37,6 куб. м м |
| Передаточное отношение дифференциала | 5,5 |
Кузов
Имеет цельнометаллическую конструкцию с задней загрузкой. Он собирается из сэндвич-панелей и обшивается оцинкованным листом изнутри и снаружи. Наружный лист имеет еще и полимерное покрытие. Пол также выполнен из сэндвич-панели и сделан усиленным.
Фургон держит температуру благодаря изотермическим свойствам применяемого материала. По заказу в зависимости от назначения и типа перевозимых грузов, кузов может быть оснащен:
- холодильной установкой;
- обогревателем;
- подъемным задним бортом, грузоподъемностью 1,5 тонны.
Кабина
У модели МАЗ 5731 она двухместная, с двумя спальными местами. Для доступа к двигателю кабина снабжена механизмом опрокидывания. Сиденья с рессорной подвеской. Панель приборов из пластика. Приборы расположены в соответствии с европейскими стандартами и отличаются информативностью. Руль оборудован гидроусилителем.
Салон аскетичен без изысков, но внутри просторно. Присутствует хорошая шумо- и виброизоляция, что создает условия для комфортной работы водителя.
Присутствует хорошая шумо- и виброизоляция, что создает условия для комфортной работы водителя.
Двигатель
В МАЗ 5731 устанавливается несколько типов. Самый распространенные это ЯМЗ-236БЕ и ЯМЗ-238ДЕ. 236. Последний – это четырехтактный V-образный шестицилиндровый дизель с турбонаддувом, мощностью – 250 л.с.
238-я модель – это уже мощный V8 для машин, работающих преимущественно с прицепами. В нем 330 сил. Также здесь присутствует турбина. Работа этого мотора сопровождается характерным свистом.
Трансмиссия
Коробка передач ЯМЗ238М – восьмиступенчатая механика с делителем. Передачи переднего хода снабжены синхронизаторами. Сцепление двухдисковое механическое. Кардан состоит из двух валов с дополнительной опорой.
Подвеска
Подвеска передней оси состоит из двух продольных полуэллиптических рессор. Задний мост опирается на основные и дополнительные рессоры. И спереди и сзади установлены стабилизаторы поперечной устойчивости.
Тормозная система
МАЗ 5731 оборудован рабочей, стояночной, запасной и вспомогательными системами торможения. Рабочая система – пневматическая, двухконтурная. Каждый контур отвечает за тормозные механизмы своей оси. Тормоза на всех колесах – барабанные. Установлена антиблокировочная система.
Рабочая система – пневматическая, двухконтурная. Каждый контур отвечает за тормозные механизмы своей оси. Тормоза на всех колесах – барабанные. Установлена антиблокировочная система.
Устройство
Кузов модели МАЗ-5336 состоит из металлической платформы с открывающимися задним и боковыми бортами. Пол кузова выполнен из дерева.
Автомобиль оснащается улучшенной трансмиссией с двухдисковым сцеплением «ЯМЗ-238Н», дополненным механическим приводом выключения с установленным пневматическим усилителем и периферийными пружинами. Грузовик имеет 4-ступенчатую механическую коробку передач с демультипликатором (общее количество передач – восемь). Карданная передача представлена двумя последовательными валами с промежуточной опорой, главная передача – двухступенчатая и разнесенная.
На МАЗ-5336 устанавливаются бездисковые колеса. Передняя подвеска автомобиля монтируется на продольных полуэллиптических рессорах со стабилизатором поперечной устойчивости и амортизаторами.
Рабочая тормозная система имеет барабанные механизмы и двухконтурный пневмопривод.
Платформа МАЗ-5336 в базовом варианте не имеет тента. Он устанавливается дополнительно. Также на шасси может монтироваться и другое оборудование.
Тягач комплектуется увеличенной кабиной, которая может откидываться посредством гидропривода. Данная возможность отличает МАЗ-5336 от более старых модификаций МАЗ. Опционально предусмотрена установка двух спальных мест, что позволяет использовать автомобиль для межрегиональных перевозок. Водителей особенно порадует главное сиденье. Оно является подрессоренным и настраивается по высоте, длине и наклону спинки и подушки. Руль также оборудован средствами регулировки.
Кабина
Кабина на «МАЗ-5336» – двухместная, подрессоренная, откидываемая вперёд посредством гидроцилиндра с ручным приводом. В базовой комплектации на «МАЗ-5336» имеются ремни безопасности, антиблокировочная система торможения ABS (НП РУП «Экран»), подрессоренное водительское кресло и предпусковой подогреватель силового агрегата.
Кресло водителя грузовика не только подрессорено, но и снабжено регулировками по длине, высоте, наклону подушки и спинки. Рулевая колонка также оборудована системой регулировок. В кабине для дальних рейсов предусмотрено два спальных места. Внутреннее пространство отличается от предшествующей модели «МАЗ»а более существенным простором и множеством дополнительных полочек. Также, кабина грузовика семейства «МАЗ-5336» оборудована задним защитным устройством; продвинутой системой шумоизоляции; электро- и пневмо- выводами.
Рулевая колонка также оборудована системой регулировок. В кабине для дальних рейсов предусмотрено два спальных места. Внутреннее пространство отличается от предшествующей модели «МАЗ»а более существенным простором и множеством дополнительных полочек. Также, кабина грузовика семейства «МАЗ-5336» оборудована задним защитным устройством; продвинутой системой шумоизоляции; электро- и пневмо- выводами.
Весовые характеристики грузовика:
- собственная масса – 8050 кг;
- общая масса – 16500 кг;
- нагрузка на переднюю ось – 6500 кг;
- нагрузка на заднюю ось – 10000 кг;
- грузоподъемность – 7700 кг;
- общая масса в составе автопоезда – 36000 кг.
Платформа модели имеет следующие габариты: длина 6080 мм, ширина – 2380 мм, высота – 2540 мм.
До 60 км/час тягач разгоняется за 50 сек, тормозной путь модели на скорости 60 км/час составляет 36,7 м. Максимальная скорость машины равняется 100 км/час.
Внутренние размеры платформы
- Длина – 6080 мм;
- Ширина – 2380 мм;
- Высота бортов -700 мм.

Модификации
Выпуск МАЗ 5336 был начат в 1988 году, и за время производства, которое продолжается и в наши дни, разработано несколько десятков различных модификаций. МАЗ 5336 используется в качестве:
- Крытых тентовых фургонов;
- Самосвалов различной грузоподъемности;
- Грузовых машин с удлинённой базой;
- Грузовиков с цельнометаллическими и сварными фургонами.
В разные годы выпускались различные версии машины, отличающиеся друг от друга мощностью двигателя, типом коробки передач, объёмов грузовой платформы наличием дополнительных опций и другими параметрами.
- МАЗ-533603-220 – двухосный грузовик с платформой площадью 10 кв.м., двигателем ЯМЗ мощностью 250 л.с.(184 кВт), допустимая полная масса автомобиля 18000 кг, допустимая масса автопоезда 36000 кг;
- МАЗ-533605-220- двухосный бортовой грузовик с платформой площадью 14,5 кв.м., двигателем ЯМЗ мощностью 330 л.с. (243 кВт), допустимая полная масса автомобиля 18750 кг, допустимая полная масса автопоезда 44000 кг.

На различных модификациях МАЗ 5336 устанавливались как атмосферные дизельные силовые установки, так и турбированные версии.
МАЗ-437121 модификации, технические характеристики — MAZ.BY
Минский автомобильный завод | МАЗ
Предназначен для городских и пригородных перевозок различных грузов.
КОНТАКТНАЯ ИНФОРМАЦИЯ:
Компания: ОАО «МАЗ» — управляющая компания холдинга «БЕЛАВТОМАЗ»
Адрес: г. Минск, ул. Социалистическая 2
Режим работы: Понедельник — Пятница: 8:00 — 16:30; Выходной: суббота, воскресенье.
- Модификации модели
Внимание! Данные сведения носят исключительно информационный характер. Информация, представленная на Сайте, не является исчерпывающей.
- МАЗ-437121-521-000
-
Экологический класс Euro 5 Колесная формула 4×2 Модель двигателя Евро-5 Коробка передач, число передач ( 6 ступ. )Объем топливного бака, в литрах 200 Мощность двигателя, кВт/л.с. 136/185 Шины 235/75R17.5 Тип кабины 4371 – малая трехместная Нагрузка на переднюю ось, кг 3800 Нагрузка на заднюю ось, кг 6500 Технически допустимая полная масса автомобиля, кг 10100 Технически допустимая полная масса автопоезда, кг — Снаряженная масса автомобиля, кг 5500 Технически допустимая грузоподъёмность, кг 4600 Оформить заявку
 )
)
- МАЗ-437121-522-000
-
Экологический класс Euro 5 Колесная формула 4×2 Модель двигателя Euro 5 Коробка передач, число передач 6 ступ. Объем топливного бака, в литрах 200 Мощность двигателя, кВт/л.с. 136/185 Шины 235/75R17.5 Тип кабины малая трёхместная Нагрузка на переднюю ось, кг 3250 Нагрузка на заднюю ось, кг 2400 Технически допустимая полная масса автомобиля, кг 10100 Технически допустимая полная масса автопоезда, кг Снаряженная масса автомобиля, кг 5650 Технически допустимая грузоподъёмность, кг 4450 Оформить заявку

- МАЗ-437121-528-000
-
Экологический класс Euro 5 Колесная формула 4×2 Модель двигателя Euro 5 Коробка передач, число передач 6 ступ. Объем топливного бака, в литрах 200 Мощность двигателя, кВт/л.с. 136/185 Шины 235/75R17.5 Тип кабины малая трёхместная Нагрузка на переднюю ось, кг 3130 Нагрузка на заднюю ось, кг 2070 Технически допустимая полная масса автомобиля, кг 10100 Технически допустимая полная масса автопоезда, кг Снаряженная масса автомобиля, кг 5200 Технически допустимая грузоподъёмность, кг 4900 Оформить заявку

- МАЗ-437121-529-000
-
Экологический класс Euro 5 Колесная формула 4×2 Модель двигателя Euro 5 Коробка передач, число передач 6 ступ. Объем топливного бака, в литрах 200 Мощность двигателя, кВт/л.с. 136/185 Шины 235/75R17.5 Тип кабины малая трёхместная Нагрузка на переднюю ось, кг 3130 Нагрузка на заднюю ось, кг 1930 Технически допустимая полная масса автомобиля, кг 10100 Технически допустимая полная масса автопоезда, кг Снаряженная масса автомобиля, кг 5050 Технически допустимая грузоподъёмность, кг 5050 Оформить заявку

- МАЗ-437121-531-000
-
Экологический класс Euro 5 Колесная формула 4×2 Модель двигателя Euro 5 Коробка передач, число передач 6 ступ. Объем топливного бака, в литрах 200 Мощность двигателя, кВт/л.с. 136/185 Шины 235/75R17.5 Тип кабины малая трёхместная Нагрузка на переднюю ось, кг 3250 Нагрузка на заднюю ось, кг 2250 Технически допустимая полная масса автомобиля, кг 10100 Технически допустимая полная масса автопоезда, кг 18000 Снаряженная масса автомобиля, кг 5500 Технически допустимая грузоподъёмность, кг 4600 Оформить заявку

- МАЗ-437121-532-000
-
Экологический класс Euro 5 Колесная формула 4×2 Модель двигателя Euro 5 Коробка передач, число передач 6 ступ. Объем топливного бака, в литрах 200 Мощность двигателя, кВт/л.с. 136/185 Шины 235 Тип кабины малая трёхместная Нагрузка на переднюю ось, кг 3250 Нагрузка на заднюю ось, кг 2400 Технически допустимая полная масса автомобиля, кг 10100 Технически допустимая полная масса автопоезда, кг 18000 Снаряженная масса автомобиля, кг Технически допустимая грузоподъёмность, кг 4450 Оформить заявку

Заявка на технику
Продукция
- Конфигуратор модели
- Спецпредложения
- Грузовая техника
- Пассажирская техника
- Прицепная техника
- Специальная техника
- Микроавтобусы
- Мероприятия по улучшению качества
- Торговый дом — запасные части
- Автотехника с пробегом (б/у)
- Автотехника МАЗ-КУПАВА шасси МАЗ
Дилеры
- Информация для дилеров
- Наши дилеры
- Как стать дилером
- Торговые дома ОАО»МАЗ»
Медиацентр
- Новости
- Элементы бренда
- Газета «АВТОЗАВОДЕЦ»
5731 — Штифт с прессовой посадкой | Mill-Max Mfg.
 Corp.
Corp.
Перейти к основному содержанию
переключить мобильное меню
Регистрация
Логин
Введите номер детали или ключевое слово
Настройка этой части
Получить техническую помощь
Узнать больше об этом продукте
Поиск 3D-модели, подождите…
Скачать 3D модель
3D-модель недоступна
Проверить наличие 3D-модели
Настройка этой части
Получить техническую помощь
Узнать больше об этом продукте
Поиск 3D-модели, пожалуйста, подождите. ..
..
Скачать 3D модель
3D-модель недоступна
Проверить наличие 3D-модели
Обзор
Описание:
Запрессовка контакта 9 печатной платы0044 Элемент крепления:
Запрессовка в сквозное отверстие без покрытия (NPTH) или изолятор
Монтажное отверстие:
0,056 дюйма (1422 мм)
Тип хвостовой части:
Паяный хвостовик
Диаметр хвостовика: 0,024 дюйма (0,610 мм)
Альтернативное крепление:
Крепление под пайку через отверстие
Альтернативное монтажное отверстие:
0,028″ (0,711 мм)
Семейство продуктов:
Печатная плата (PCB), контакт
Сопряжения с:
Найдите места для спаривания
Технические характеристики
Номинальный ток:
Специальное приложение
Диапазон рабочих температур:
— 55/+125°C
Настройка этой части
Получить техническую помощь
Узнать больше об этом продукте
Поиск 3D-модели, подождите. ..
..
Скачать 3D модель
3D-модель недоступна
Проверить наличие 3D-модели
Обзор
Описание:
Штифт печатной платы с запрессовкой
Монтажная характеристика:
Запрессовка в сквозное отверстие без покрытия (NPTH) или изолятор
Монтажное отверстие:
0,056 дюйма (1422 мм)
Тип хвоста:
Паяный хвостовик
Диаметр хвостовика: 0,024 дюйма (0,610 мм)
Альтернативное крепление:
Крепление под пайку через отверстие
Альтернативное монтажное отверстие:
0,028″ (0,711 мм)
Семейство продуктов:
Вывод печатной платы (PCB)
Сочетается с:
Найдите места для спаривания
Технические характеристики
Номинальный ток:
Для конкретного применения
Диапазон рабочих температур:
— 55/+125°C
Продукты
фильтр
Загрузка продуктов
| Деталь № | Приоритет | В наличии | В наличии Сортировка | Наружная обшивка | Покрытие контактов | ROHS | Контактный номер | Материал корпуса | Варианты упаковки | Кол-во в катушке | Жизненный цикл | Примечание для запроса | Запрос коммерческого предложения/образец | Технический паспорт | Изображение |
|---|
Отображение XX из XX результатов
Теги
npthpress-fitпечатная плата pinpcb pin
Недавно просмотренные
Интеграция метаболизма липидов, продукции феромонов и восприятия ядерным фактором бесплодия и гепатоцитов 4
1. Халлаф М. А.
Халлаф М. А.
и др., Крупномасштабная характеристика коммуникационных систем половых феромонов у дрозофилы. Связь с природой
12, (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Вятт Т. Д., Феромоны и смеси сигнатур: определение видовых сигналов и переменных сигналов для идентификации как у беспозвоночных, так и у позвоночных. Журнал сравнительной физиологии А
196, 685–700 (2010). [PubMed] [Google Scholar]
3. Тулсон Э. С., Влияние температуры выращивания на проницаемость кутикулы и состав эпикутикулярных липидов у Drosophila pseudoobscura. Журнал экспериментальной зоологии
222, 249–253 (1982). [Google Scholar]
4. Локки К. Х. Липиды кутикулы насекомых: происхождение, состав и функции. Сравнительная биохимия и физиология Часть B: Сравнительная биохимия
89, 595–645 (1988). [Google Scholar]
5. Rouault J.-D., Marican C., Wicker-Thomas C., Jallon J.-M., in Drosophila melanogaster, Drosophila simulans: такие похожие, такие разные. (Спрингер, 2004), стр. 195–212. [PubMed] [Google Scholar]
195–212. [PubMed] [Google Scholar]
6. Оцу Т., Кимура М. Т., Катагири С., Как виды дрозофил приобретают холодостойкость: качественные изменения фосфолипидов. Европейский журнал биохимии
252, 608–611 (1998). [PubMed] [Google Scholar]
7. Хоффманн А. А., Халлас Р., Синклер К., Митровски П., Уровни изменчивости стрессоустойчивости дрозофилы среди штаммов, местных популяций и географических регионов: закономерности высыхания, голодания, холодостойкость и связанные с ней признаки. Эволюция
55, 1621–1630 (2001). [PubMed] [Академия Google]
8. Кобб М., Джаллон Дж.-М., Феромоны, распознавание партнера и стимуляция ухаживания в подгруппе видов Drosophila melanogaster. Поведение животных
39, 1058–1067 (1990). [Google Scholar]
9. Койн Дж. А., Криттенден А. П., Мах К., Генетика различий феромонов, способствующих репродуктивной изоляции у дрозофилы. Наука
265, 1461–1464 (1994). [PubMed] [Google Scholar]
10. Этгес В. Дж., Аренс М. А., Изоляция перед спариванием определяется субстратами для выращивания личинок у кактофильной Drosophila mojavensis. V. Глубокая географическая изменчивость эпикутикулярных углеводородов среди изолированных популяций. Американский натуралист
V. Глубокая географическая изменчивость эпикутикулярных углеводородов среди изолированных популяций. Американский натуралист
158, 585–598 (2001). [PubMed] [Google Scholar]
11. Ferveur J.-F. Кутикулярные углеводороды: их эволюция и роль в феромональной коммуникации дрозофилы. Генетика поведения
35, 279–295 (2005). [PubMed] [Google Scholar]
12. Вятт Т. Д., Феромоны и поведение животных. (Издательство Кембриджского университета, Кембридж, 2003 г.), том. 626. [Google Scholar]
13. Makki R., Cinnamon E., Gould A.P., Развитие и функции эноцитов. Ежегодный обзор энтомологии
59, 405–425 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Уикер-Томас К.
и др., Гибкое происхождение предшественников углеводородов/феромонов у Drosophila melanogaster [S]. Журнал исследований липидов
56, 2094–2101 (2015). [PMC free article] [PubMed] [Google Scholar]
15. Fan Y., Zurek L., Dykstra M.J., Schal C. Синтез углеводородов ферментативно диссоциированными эноцитами брюшного покрова немецкого таракана, Blattella germanica.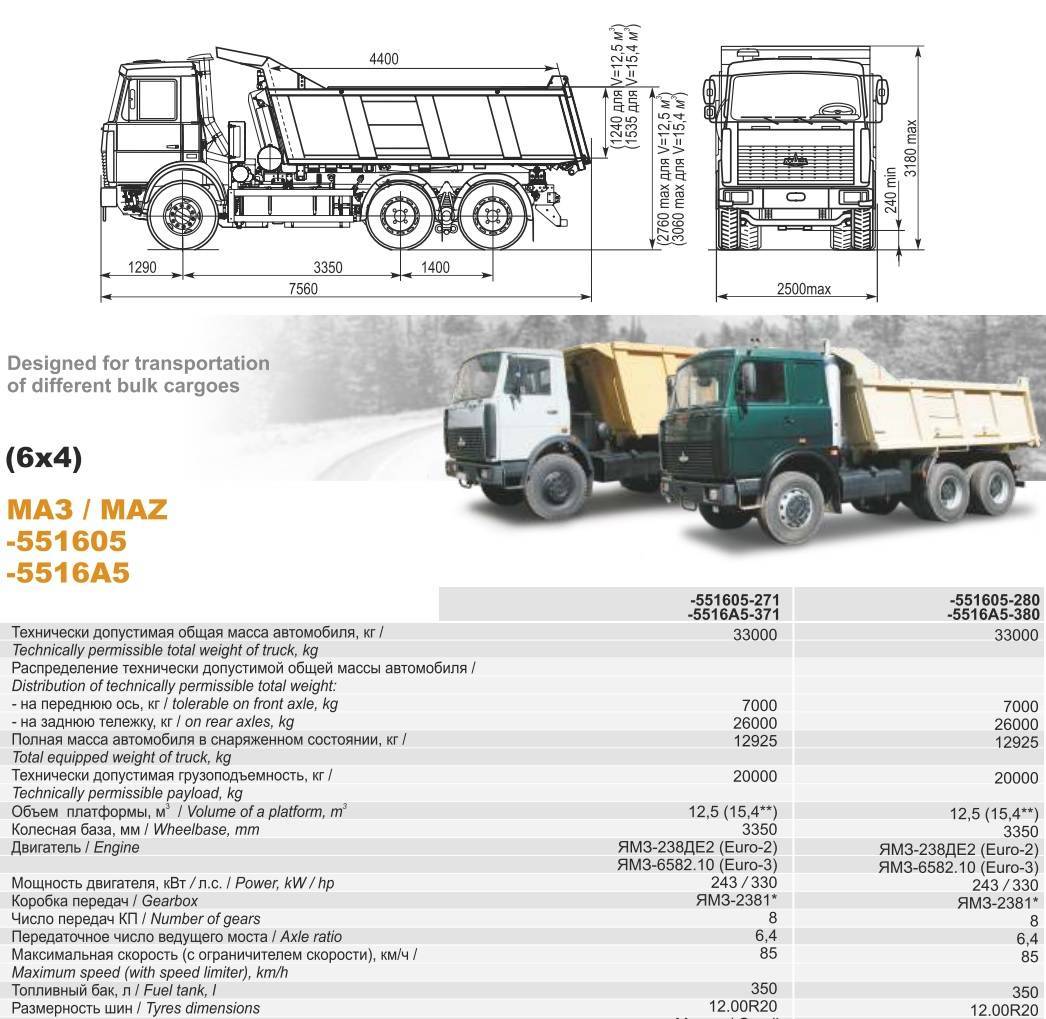 Naturwissenschaften
Naturwissenschaften
90, 121–126 (2003). [PubMed] [Google Scholar]
16. Чанг Х.
и др., Характеристика генов цитохрома P450 Drosophila melanogaster. Труды Национальной академии наук
106, 5731–5736 (2009 г.)). [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Gutierrez E., Wiggins D., Fielding B., Gould A.P., Специализированные гепатоцитоподобные клетки регулируют метаболизм липидов дрозофилы. Природа
445, 275–280 (2007). [PubMed] [Google Scholar]
18. Кунте А. С., Мэтьюз К. А., Роусон Р. Б., Ауксотрофия жирных кислот у личинок дрозофилы, лишенных SREBP. Метаболизм клеток
3, 439–448 (2006). [PubMed] [Google Scholar]
19. Lycett G.
et al., Anopheles gambiae P450 редуктаза в высокой степени экспрессируется в эноцитах, и нокдаун in vivo увеличивает чувствительность к перметрину. Молекулярная биология насекомых
15, 321–327 (2006). [PubMed] [Академия Google]
20. Мартиньш Г. Ф.
и др., Изучение транскриптома эноцитов куколок Aedes aegypti. Памяти Института Освальдо Круза
106, 308–315 (2011). [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
21. Парви Ж.-П.
et al., Ацетил-КоА-карбоксилаза Drosophila melanogaster поддерживает удаленный сигнал, зависящий от жирных кислот, для гидроизоляции дыхательной системы. (2012). [Бесплатная статья PMC] [PubMed]
22. Хуан К.
и др., Транслатомное профилирование RiboTag эноцитов дрозофилы при старении и индуцированном окислительном стрессе. Геномика BMC
20, 50 (2019). [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Ховард Р. В., Бломквист Г. Дж., Экологические, поведенческие и биохимические аспекты углеводородов насекомых. Ежегодный обзор энтомологии
50, 371–393 (2005). [PubMed] [Google Scholar]
24. Ширанги Т. Р., Дюфур Х. Д., Уильямс Т. М., Кэрролл С. Б., Быстрая эволюция экспрессии фермента, продуцирующего половые феромоны, у дрозофилы. биология PLoS
7, e1000168 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Чертемпс Т.
et al., Экспрессируемая элонгаза, ориентированная на самок, участвует в биосинтезе длинноцепочечных углеводородов и ухаживании у Drosophila melanogaster. Труды Национальной академии наук
Труды Национальной академии наук
104, 4273–4278 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Chertemps T., Duportets L., Labeur C., Ueyama M., Wicker-Thomas C., Женский специфический ген десатуразы, ответственный за биосинтез диеновых углеводородов и поведение при ухаживании у Drosophila melanogaster. Насекомое Мол Биол
15, 465–473 (2006). [PubMed] [Google Scholar]
27. Holze H., Schrader L., Buellesbach J., Успехи в расшифровке генетической основы биосинтеза и изменчивости кутикулярных углеводородов насекомых. Наследственность
126, 219–234 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Kimura K.-i., Hachiya T., Koganezawa M., Tazawa T., Yamamoto D., Бесплодные и двуполые координируют создание мужских нейронов, которые могут инициировать ухаживание. Нейрон
59, 759–769 (2008). [PubMed] [Google Scholar]
29. Kohatsu S., Koganezawa M., Yamamoto D., Женский контакт активирует мужские интернейроны, которые запускают стереотипное ухаживание у дрозофилы. Нейрон
Нейрон
69, 498–508 (2011). [PubMed] [Google Scholar]
30. Rideout E. J., Billeter J.-C., Goodwin S. F. Гены определения пола бесплодия и двойного пола определяют нейронный субстрат, необходимый для песни ухаживания. Текущая биология
17, 1473–1478 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Фон Филипсборн А.С.
и др., Нейрональный контроль песни ухаживания дрозофилы. Нейрон
69, 509–522 (2011). [PubMed] [Google Scholar]
32. Асахина К.
et al., Тахикинин-экспрессирующие нейроны контролируют специфическое для самцов агрессивное возбуждение у дрозофилы. Клетка
156, 221–235 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Ishii K., Wohl M., DeSouza A., Asahina K., Гены, определяющие пол, отчетливо регулируют способность к ухаживанию и целевые предпочтения через половые диморфные нейроны. Элиф
9, e52701 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Коганезава М., К.-и. Кимура, Д. Ямамото, Нервная схема, которая функционирует как переключатель ухаживания против агрессии у самцов дрозофилы. Текущая биология
Текущая биология
26, 1395–1403 (2016). [PubMed] [Google Scholar]
35. Чен Д.
и др., Генетические и нейронные механизмы, управляющие специфическим для пола взаимодействием между сном и половым поведением у дрозофилы. Связь с природой
8, 1–14 (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Усуи-Аоки К.
и др., Формирование мужских мышц у самок дрозофилы путем эктопической бесплодной экспрессии. Природа клеточной биологии
2, 500–506 (2000). [PubMed] [Google Scholar]
37. Ryner L.C.
и др., Контроль мужского сексуального поведения и сексуальной ориентации у дрозофилы бесплодным геном. Клетка
87, 1079–1089 (1996). [PubMed] [Google Scholar]
38. Ананд А.
и др., Молекулярно-генетическое вскрытие половых и жизненных функций гена определения пола Drosophila melanogaster безрезультатно. Генетика
158, 1569–1595 (2001). [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Song H.-J.
et al., Бесплодный ген необходим для правильного формирования аксональных трактов в эмбриональной центральной нервной системе дрозофилы. Генетика
Генетика
162, 1703–1724 (2002). [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Zhou H., Whitworth C., Pozmanter C., Neville M.C., Van Doren M., Doublesex регулирует бесплодную экспрессию, чтобы способствовать половому диморфизму ниши стволовых клеток гонады . Генетика PLoS
17, е1009468 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Сато К., Ямамото Д., Взаимоисключающая экспрессия половых и неполовых бесплодных генных продуктов в центральной нервной системе дрозофилы. Паттерны экспрессии генов
43, 119232 (2022). [PubMed] [Google Scholar]
42. Аллен С. Э.
и др., Универсальный мозаичный анализ, опосредованный CRISPR/Cas9, с помощью гРНК-индуцированного кроссинговера для немодифицированных геномов. биология PLoS
19, e3001061 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Гилл К. С., Мутация, вызывающая аномальное ухаживание и брачное поведение у самцов Drosophila melanogaster. Являюсь. Зоол.
3, 507 (1963). [Google Scholar]
44. Виллелла А.
Виллелла А.
и др., Расширенные репродуктивные роли гена бесплодия у Drosophila melanogaster, выявленные с помощью поведенческого анализа новых мутантов fru. Генетика
147, 1107–1130 (1997). [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Биллетер Дж.-К., Аталлах Дж., Крупп Дж.Дж., Миллар Дж.Г., Левин Дж.Д., Специализированные клетки отмечают половую и видовую идентичность Drosophila melanogaster. Природа
461, 987–991 (2009). [PubMed] [Google Scholar]
46. Амрейн Х. Восприятие феромонов и поведение у дрозофилы. Текущее мнение в нейробиологии
14, 435–442 (2004). [PubMed] [Google Scholar]
47. Могильнер А., Эдельштейн-Кешет Л., Бент Л., Спирос А. Взаимные взаимодействия, потенциалы и индивидуальная дистанция в социальной совокупности. Журнал математической биологии
47, 353–389 (2003). [PubMed] [Google Scholar]
48. Greenspan R.J., Ferveur J.-F., Ухаживание за дрозофилой. Ежегодный обзор генетики
34, 205–232 (2000). [PubMed] [Академия Google]
49. Лакай Ф.
Лакай Ф.
et al., Ингибирующий половой феромон имеет горький вкус для самцов дрозофилы. PloS один
2, e661 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Snellings Y.
и др., Роль кутикулярных углеводородов в распознавании партнеров у Drosophila suzukii. Научные отчеты
8, 4996 (2018). [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Джаллон Дж. М., Дэвид Дж. Р., Вариации кутикулярных углеводородов среди восьми видов подгруппы Drosophila melanogaster. Эволюция
41, 294–302 (1987). [PubMed] [Google Scholar]
52. Цю Ю.
et al., Специфичная для насекомых окислительная декарбонилаза P450 для биосинтеза кутикулярных углеводородов. Труды Национальной академии наук
109, 14858–14863 (2012). [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Billeter J.-C.
и др., Изоформ-специфический контроль дифференцировки и поведения мужских нейронов у дрозофилы с помощью бесплодного гена. Текущая биология
16, 1063–1076 (2006). [PubMed] [Google Scholar]
54. Чен Дж.
Чен Дж.
et al., бесплодно настраивает функциональную гибкость схемы ухаживания во время развития. электронная жизнь
10, е59224 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Стокингер П., Квициани Д., Роткопф С., Тириан Л., Диксон Б. Дж., Нейронная схема, регулирующая поведение самцов дрозофилы при ухаживании. Клетка
121, 795–807 (2005). [PubMed] [Google Scholar]
56. Kimura K.-I., Ote M., Tazawa T., Yamamoto D., Fruitless определяет половые диморфные нейронные схемы в мозге дрозофилы. Природа
438, 229–233 (2005). [PubMed] [Google Scholar]
57. Бломквист Г. Дж., Баньер А.-Г. Углеводороды насекомых: биология, биохимия и химическая экология. (Издательство Кембриджского университета, 2010 г.). [Академия Google]
58. Корица E.
et al., Drosophila Spidey/Kar регулирует рост эноцитов посредством передачи сигналов PI3-киназы. Генетика PLoS
12, e1006154 (2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Storelli G., Nam H.-J., Simcox J. , Villanueva C.J., Thummel C.S., Drosophila HNF4 управляет переключением в метаболизме липидов, которое поддерживает переход к Совершеннолетие. Ячейка развития
, Villanueva C.J., Thummel C.S., Drosophila HNF4 управляет переключением в метаболизме липидов, которое поддерживает переход к Совершеннолетие. Ячейка развития
48, 200–214.e206 (2019). [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Паланкер Л., Теннессен Дж. М., Лам Г., Таммел С. С., Drosophila HNF4 регулирует мобилизацию липидов и β-окисление. Метаболизм клеток
9, 228–239 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Палу Р. А., Туммел К. С., Sir2 действует через ядерный фактор 4 гепатоцитов для поддержания передачи сигналов инсулина и метаболического гомеостаза у дрозофилы. Генетика PLoS
12, e1005978 (2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Pei X.-J.
и др. Модуляция удлинения жирных кислот у тараканов поддерживает половой диморфизм углеводородов и привлекательность самок. биология PLoS
19, e3001330 (2021). [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Чен Н.
и др., Один ген интегрирует регуляторы пола и гормонов в сексуальную привлекательность. Экология природы и эволюция (2022). [PubMed] [Google Scholar]
Экология природы и эволюция (2022). [PubMed] [Google Scholar]
64. Fernández M. d. л. П.
et al., Феромональные и поведенческие сигналы вызывают агрессию самца к самке у дрозофилы. биология PLoS
8, e1000541 (2010). [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Паркер Д. Дж., Гардинер А., Невилл М. К., Ричи М. Г., Гудвин С. Ф., Эволюция новизны в законсервированных генах; свидетельство положительного отбора в бесплодном гене дрозофилы локализовано в альтернативно сплайсированных экзонах. Наследственность
112, 300–306 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Дорнан А. Дж., Гейли Д. А., Гудвин С. Ф., Нацеливание энхансерной ловушки GAL4 на ген определения пола дрозофилы безрезультатно. генезис
42, 236–246 (2005). [PubMed] [Google Scholar]
67. Чоудхури Т., Калхун Р. М., Брух К., Мёринг А. Дж., Бесплодный ген влияет на восприимчивость самок и изоляцию видов. Труды Королевского общества B
287, 20192765 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
68. Lee G.
Lee G.
и др., Пространственные, временные и половые диморфные паттерны экспрессии бесплодного гена в центральной нервной системе дрозофилы. Журнал нейробиологии
43, 404–426 (2000). [PubMed] [Академия Google]
69. Нг С. Х.
et al., Эволюция феромонов и сексуальное поведение дрозофилы формируются за счет сенсорной эксплуатации самцами других самцов. Труды Национальной академии наук
111, 3056–3061 (2014). [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Деккер Т.
et al., Потеря феромона дрозофилы меняет его роль в половом общении у дрозофилы suzukii. Труды Королевского общества B: биологические науки
282, 2014–3018 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
71. Джозеф Р. М., Карлсон Дж. Р., Хеморецепторы дрозофилы: молекулярный интерфейс между химическим миром и мозгом. Тенденции Жене
31, 683–695 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
72. Маноли Д. С.
et al., Специфические для самцов бесплодные определяют нейронные субстраты поведения ухаживания дрозофилы. Природа
Природа
436, 395–400 (2005). [PubMed] [Google Scholar]
73. Родригес В., Хаммель Т., Развитие обонятельной системы дрозофилы. Adv Exp Мед Биол
628, 82–101 (2008). [PubMed] [Академия Google]
74. Сети С.
и др., Социальный контекст усиливает гормональную модуляцию обнаружения феромонов у дрозофилы. Текущая биология
29, 3887–3898.e3884 (2019). [Бесплатная статья PMC] [PubMed] [Google Scholar]
75. Zhang Y., Ng R., Neville M.C., Goodwin S.F., Su C.-Y., Различная роль и синергетическая функция изоформ FruM в нейронах обонятельных рецепторов дрозофилы . Отчеты о ячейках
33, 108516 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
76. Ван Дж.
и др., Регуляция дифференцировки нервных стволовых клеток факторами транскрипции HNF4-1 и MAZ-1. Молекулярная нейробиология
47, 228–240 (2013). [PubMed] [Академия Google]
77. Laranjeira A., Schulz J., Dotti C.G., Гены, связанные с β-окислением жирных кислот, играют роль в функциональном снижении мозга дрозофилы с возрастом. PLoS Один
PLoS Один
11, e0161143 (2016). [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Лессинг Д., Бонини Н. М., Поддержание мозга: понимание нейродегенерации человека от мутантов Drosophila melanogaster. Природа Обзоры Генетика
10, 359–370 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Jallon J.-M., Lauge G., Orssaud L., Antony C., Женские феромоны у Drosophila melanogaster контролируются двуполым локусом. Генетические исследования
51, 17–22 (1988). [Google Scholar]
80. Томпкинс Л., МакРоберт С. П., Поведенческие и феромональные фенотипы, связанные с экспрессией мутаций с потерей функции в летальном для пола гене Drosophila melanogaster. Журнал нейрогенетики
9, 219–226 (1995). [PubMed] [Google Scholar]
81. Томпкинс Л., МакРоберт С., Регуляция поведенческих и феромональных аспектов определения пола у Drosophila melanogaster с помощью сексуально летального гена. Генетика
123, 535–541 (1989). [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Уотербери Дж. А., Джексон Л. Л., Шедл П., Анализ двуполого женского белка у Drosophila melanogaster: роль в половой дифференциации и поведении и зависимости от интерсексуалов. Генетика
Уотербери Дж. А., Джексон Л. Л., Шедл П., Анализ двуполого женского белка у Drosophila melanogaster: роль в половой дифференциации и поведении и зависимости от интерсексуалов. Генетика
152, 1653–1667 (1999). [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Ferveur J.-F.
и др., Генетическая феминизация феромонов и ее поведенческие последствия у самцов дрозофилы. Наука
276, 1555–1558 (1997). [PubMed] [Google Scholar]
84. Шарма М. Д., Хант Дж., Хоскен Д. Дж., Антагонистические реакции на естественный и половой отбор и специфическая для пола эволюция кутикулярных углеводородов у Drosophila simulans. Эволюция: Международный журнал органической эволюции
66, 665–677 (2012). [PubMed] [Академия Google]
85. Wicker C., Jallon J.-M. Влияние яичников и экдистероидов на биосинтез феромонов у Drosophila melanogaster (Diptera: Drosophilidae). Европейский журнал энтомологии
92, 197–197 (1995). [Google Scholar]
86. Билен Дж., Аталлах Дж., Азанчи Р., Левин Дж. Д., Риддифорд Л. М. Регулирование начала спаривания самок и производства половых феромонов ювенильным гормоном у Drosophila melanogaster. Труды Национальной академии наук
М. Регулирование начала спаривания самок и производства половых феромонов ювенильным гормоном у Drosophila melanogaster. Труды Национальной академии наук
110, 18321–18326 (2013). [Бесплатная статья PMC] [PubMed] [Google Scholar]
87. Далтон Дж. Э., Лебо М. С., Сандерс Л. Э., Сан Ф., Арбайтман М. Н., Экдизоновые рецепторы действуют в бесплодных экспрессирующих нейронах для опосредования ухаживания дрозофилы. Текущая биология
19, 1447–1452 (2009). [Бесплатная статья PMC] [PubMed] [Google Scholar]
88. Чжан Б., Сато К., Ямамото Д., Передача сигналов экдизона регулирует спецификацию нейронов с мужскими нейритами у дрозофилы. Биология открыта
7, био029744 (2018). [Бесплатная статья PMC] [PubMed] [Google Scholar]
89. Чжао С.
и др., Перепрограммирование регулятора ухаживания на основе хроматина путем одновременного восприятия феромонов и передачи сигналов гормонами. Научные достижения
6, eaba6913 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
90. Wang X.
и др., Взаимодействие гормонов и рецепторов в регуляции метаболизма липидов у комаров. Труды Национальной академии наук
114, E2709–E2718 (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
91. Салливан А. А., Таммел К. С., Временные профили экспрессии генов ядерных рецепторов показывают координированные транскрипционные ответы во время развития дрозофилы. Молекулярная эндокринология
17, 2125–2137 (2003). [PubMed] [Академия Google]
92. Хуанг К.
et al., Нарушение импорта пероксисом в эноцитах дрозофилы вызывает сердечную дисфункцию, индуцируя upd3 в качестве пероксикина. Связь с природой
11, 2943 (2020). [Бесплатная статья PMC] [PubMed] [Google Scholar]
93. Двек Х.К.М.
и др., Феромоны, опосредующие копуляцию и влечение у дрозофилы. Труды Национальной академии наук
112, E2829–E2835 (2015). [Статья бесплатно PMC] [PubMed] [Google Scholar]
94. Everaerts C., Farine J.-P., Cobb M., Ferveur J.-F., Новый взгляд на кутикулярные углеводороды дрозофилы: статус спаривания изменяет кутикулярные профили.
